Глава 6. СИСТЕМА КРОВИ
Внутренняя среда организма представлена тканевой (интерстициальной)
жидкостью, лимфой и кровью, состав и свойства которых теснейшим образом связаны
между собой. Однако истинной внутренней средой организма является тканевая
жидкость, так как лишь она контактирует с клетками организма. Кровь же,
соприкасаясь непосредственно с эндокардом и эндотелием сосудов, обеспечивает их
жизнедеятельность и лишь косвенно через тканевую жидкость вмешивается в работу
всех без исключения органов и тканей. Через сосудистую стенку в кровоток
транспортируются гормоны и различные биологически активные соединения.
Основной составной частью тканевой жидкости, лимфы и крови является вода. В
организме человека вода составляет 75% от массы тела. Для человека массой тела
70 кг
тканевая жидкость и лимфа составляют до 30% (20—21 л), внутриклеточная жидкость
— 40% (27—29 л) и плазма — около 5% (2,8—3,0 л).
Между кровью и тканевой жидкостью происходят постоянный обмен веществ и
транспорт воды, несущей растворенные в ней продукты обмена, гормоны, газы,
биологически активные вещества. Следовательно, внутренняя среда организма
представляет собой единую систему гуморального транспорта, включающую общее
кровообращение и движение в последовательной цепи: кровь — тканевая жидкость —
ткань (клетка) — тканевая жидкость — лимфа — кровь.
Из этой простой схемы видно, насколько тесно связан состав крови не только с
тканевой жидкостью, но и с лимфой. В организме важная роль отводится
лимфатической системе, начало которой составляют лимфатические капилляры,
дренирующие все тканевые пространства и сливающиеся в более крупные сосуды. По
ходу лимфатических сосудов располагаются лимфатические узлы, при прохождении
которых изменяется состав лимфы и она обогащается лимфоцитами. Свойства лимфы,
как и тканевой жидкости, во многом определяются органом, от которого она
оттекает. После приема пищи состав лимфы резко изменяется, так как в нее
всасываются жиры, углеводы и даже белки.
Следует заметить, что внутриклеточная жидкость, плазма крови, тканевая
жидкость и лимфа имеют различный состав, что в значительной степени
определяет интенсивность водного, ионного и электролитного обмена, катионов,
анионов и продуктов метаболизма между кровью, тканевой жидкостью и клетками.
Еще в 1878
г. К. Бернар писал, что «... поддержание постоянства
условий жизни в нашей внутренней среде — необходимый элемент свободной и
независимой жизни». Это положение легло в основу учения о гомеостазе, создателем
которого является американский физиолог У. Кеннон (см. раздел 1.3). Между тем в
основе представлений о гомеостазе лежат динамические процессы, ибо
«постоянство внутренней среды организма» редко бывает постоянным. Под
влиянием внешних воздействий и сдвигов, происходящих в самом организме, состав
тканевой жидкости, лимфы и крови на короткое время может изменяться в широких
пределах, однако благодаря регуляторным воздействиям, осуществляемым нервной
системой и гуморальными факторами, сравнительно быстро возвращается к
норме. Более длительные сдвиги в гомеостазе не только сопровождают развитие
патологического процесса, но и зачастую несовместимы с жизнью.
Говоря о внутренней среде организма в этом разделе, мы коснемся лишь
физиологии системы крови. Что же касается функций тканевой жидкости и лимфы, то
они будут рассмотрены в специальной главе учебника.
6.1. ПОНЯТИЕ О СИСТЕМЕ КРОВИ
Отечественный клиницист Г. Ф. Ланг считал, что в систему крови входят кровь,
органы кроветворения и кроверазрушения, а также аппарат регуляции. Кровь как
ткань обладает следующими особенностями: 1) все ее составные части
образуются за пределами сосудистого русла; 2) межклеточное вещество ткани
является жидким; 3) основная часть крови находится в постоянном движении.
Кровь животных заключена в систему замкнутых трубок — кровеносных сосудов.
Кровь состоит из жидкой части — плазмы и форменных элементов — эритроцитов,
лейкоцитов и тромбоцитов. У взрослого человека форменные элементы крови
составляют около 40—48%, а плазма — 52—60%. Это соотношение получило название
гематокритного числа (от греч. haima — кровь, kritos — показатель). В
практической деятельности для характеристики гематокритного числа указывается
лишь показатель плотной части крови.
6.1.1. Основные функции
крови
Основными функциями крови являются транспортная, защитная и регуляторная,
остальные функции, приписываемые системе крови, являются лишь производными
основных ее функций. Все три основные функции крови связаны между собой и
неотделимы друг от друга.
Транспортная функция. Кровь переносит необходимые для
жизнедеятельности органов и тканей различные вещества, газы и продукты
обмена. Транспортная функция осуществляется как плазмой, так и форменными
элементами. Последние могут переносить все вещества, входящие в состав крови.
Многие из них переносятся в неизмененном виде, другие вступают в нестойкие
соединения с различными белками. Благодаря транспорту осуществляется
дыхательная функция крови. Кровь осуществляет перенос гормонов,
питательных веществ, продуктов обмена, ферментов, различных биологически
активных веществ, солей, кислот, щелочей, катионов, анионов, микроэлементов и
др. С транспортом связана и экскреторная функция крови — выделение из организма
метаболитов, отслуживших свой срок или находящихся в данный момент в избытке
веществ.
Защитные функции. Чрезвычайно разнообразны. С наличием в крови
лейкоцитов связана специфическая (иммунитет) и неспецифическая (главным
образом фагоцитоз) защита организма. В составе крови содержатся все
компоненты так называемой системы комплемента, играющей важную роль, как в
специфической, так и неспецифической защите. К защитным функциям относится
сохранение циркулирующей крови в жидком состоянии и остановка кровотечения
(гемостаз) в случае нарушения целостности сосудов.
Гуморальная регуляция деятельности организма. В первую очередь связана
с поступлением в циркулирующую кровь гормонов, биологически активных веществ и
продуктов обмена. Благодаря регуляторной функции крови осуществляется сохранение
постоянства внутренней среды организма, водного и солевого баланса тканей и
температуры тела, контроль за интенсивностью обменных процессов, регуляция
гемопоэза и других физиологических функций.
6.1.2. Количество крови в
организме
У человека кровь составляет 6—8% от массы тела, т. е. в среднем 5—6 л.
Определение количества крови в организме заключается в следующем: в кровь
вводят нейтральную краску, радиоактивные изотопы или коллоидный раствор и
через определенное время, когда вводимый маркер равномерно распределится,
определяют его концентрацию. Зная количество введенного вещества, легко
рассчитать количество крови в организме. При этом следует учитывать,
распределяется ли вводимый субстрат в плазме или полностью проникает в
эритроциты. В дальнейшем определяют гематокритное число, после чего производят
расчет общего количества крови в организме.
6.1.3. Состав плазмы крови
Плазма представляет собой жидкую часть крови желтоватого цвета, слегка
опалесцирующую, в состав которой входят различные соли (электролиты), белки,
липиды, углеводы, продукты обмена, гормоны, ферменты, витамины и
растворенные в ней газы (табл. 6.1).
Таблица 6.1 Химический состав плазмы крови
|
Компоненты плазмы |
Содержание, % |
Компоненты плазмы |
Содержание, % |
|
Вода |
90,5 |
Натрий |
0,3 |
|
Белки |
8 |
Калий |
0,02 |
|
Липиды |
0,3 |
Кальций |
0,012 |
|
Нейтральный жир |
0,2 |
Магний |
0,0002 |
|
Глюкоза |
0,12 |
Хлор |
0,35 |
|
Мочевина |
0,03 |
Гидрокарбонат |
0,16 |
|
Мочевая кислота |
0,004 |
Фосфат |
0,03 |
|
Креатин |
0,006 |
Сульфат |
0,02 |
|
Аминокислоты |
0,008 |
|
|
Состав плазмы отличается лишь относительным постоянством и во многом зависит
от приема пищи, воды и солей. В то же время концентрация глюкозы, белков, всех
катионов, хлора и гидрокарбонатов удерживается в плазме на довольно
постоянном уровне и лишь на короткое время может выходить за пределы нормы.
Значительные отклонения этих показателей от средних величин на длительное
время приводят к тяжелейшим последствиям для организма, зачастую
несовместимым с жизнью. Содержание же других составных элементов плазмы —
фосфатов, мочевины, мочевой кислоты, нейтрального жира может варьировать в
довольно широких пределах, не вызывая расстройств функции организма. В общей
сложности минеральные вещества плазмы составляют около 0,9%. Содержание глюкозы
в крови 4,5—6,5 ммоль/л.
Растворы, имеющие одинаковое с кровью осмотическое давление, получили
название изотонических, или физиологических. К таким
растворам для теплокровных животных и человека относится 0,9% раствор натрия
хлорида и 5% раствор глюкозы. Растворы, имеющие большее осмотическое давление,
чем кровь, называются гипертоническими, а меньшее —
гипотоническими.
Для обеспечения жизнедеятельности изолированных органов и тканей, а также при
кровопотере используют растворы, близкие по ионному составу к плазме крови
(табл. 6.2).
Таблица 6.2. Состав растворов Рингера—Локка и Тироде для теплокровных
животных
|
Название раствора |
Ингредиенты, г/л воды |
|
NaCI |
KCI |
CaCl2 |
NaHCO3 |
MgCl2 |
NaH2PO4 |
глюкоза |
|
Рингера— Локка
|
9,0 |
0,42 |
0,24 |
0,15 |
-- |
-- |
1,0 |
|
Тироде |
8,0 |
0,2 |
0,2 |
1,0 |
0,1 |
0,05 |
1,0 |
Из-за отсутствия коллоидов (белков) растворы Рингера—Локка и Тироде
неспособны на длительное время задерживать воду в крови — вода быстро выводится
почками и переходит в ткани. Поэтому в клинической практике эти растворы
применяются в качестве кровезамещающих лишь в случаях, когда отсутствуют
коллоидные растворы, способные на длительное время восполнить недостаток
жидкости в сосудистом русле.
Важнейшей составной частью плазмы являются белки, содержание которых
составляет 7—8% от массы плазмы. Белки плазмы — альбумины, глобулины и
фибриноген. К альбуминам относятся белки с относительно малой молекулярной
массой (около 70 000), их 4— 5%, к глобулинам — крупномолекулярные белки
(молекулярная масса до 450 000) — количество их доходит до 3%. На долю
глобулярного белка фибриногена (молекулярная масса 340 000) приходится
0,2—0,4%. С помощью метода электрофореза, основанного на различной скорости
движения белков в электрическом поле, глобулины могут быть разделены на
α1-, α2- и γ-глобулины.
Функции белков плазмы крови весьма разнообразны: белки обеспечивают
онкотическое давление крови, от которого в значительной степени зависит обмен
воды и растворенных в ней веществ между кровью и тканевой жидкостью; регулируют
рН крови благодаря наличию буферных свойств; влияют на вязкость крови и плазмы,
что чрезвычайно важно для поддержания нормального уровня кровяного
давления, обеспечивают гуморальный иммунитет, ибо являются антителами
(иммуноглобулинами); принимают участие в свертывании крови; способствуют
сохранению жидкого состояния крови, так как входят в состав противосвертывающих
веществ, именуемых естественными антикоагулянтами; служат переносчиками рада
гормонов, липидов, минеральных веществ и др.; обеспечивают процессы
репарации, роста и развития различных клеток организма.
6.1.4. Физико-химические свойства
крови
Цвет крови. Определяется наличием в эритроцитах особого белка —
гемоглобина. Артериальная кровь характеризуется ярко-красной окраской, что
зависит от содержания в ней гемоглобина, насыщенного кислородом
(оксигемоглобин). Венозная кровь имеет темно-красную с синеватым оттенком
окраску, что объясняется наличием в ней не только окисленного, но и
восстановленного гемоглобина. Чем активнее орган и чем больше отдал кислорода
тканям гемоглобин, тем более темной выглядит венозная кровь.
Относительная плотность крови. Колеблется от 1,058 до 1,062 и зависит
преимущественно от содержания эритроцитов. Относительная плотность плазмы
крови в основном определяется концентрацией белков и составляет
1,029—1,032.
Вязкость крови. Определяется по отношению к вязкости воды и
соответствует 4,5—5,0. Вязкость крови зависит главным образом от содержания
эритроцитов и в меньшей степени от белков плазмы. Вязкость венозной крови
несколько больше, чем артериальной, что обусловлено поступлением в эритроциты
СО2, благодаря чему незначительно увеличивается их размер.
Вязкость крови возрастает при опорожнении депо крови, содержащей большее число
эритроцитов. Вязкость плазмы не превышает 1,8—2,2. При обильном белковом
питании вязкость плазмы, а, следовательно, и крови может повышаться.
Осмотическое давление крови. Осмотическим давлением называется
сила, которая заставляет переходить растворитель (для крови это вода) через
полупроницаемую мембрану из менее в более концентрированный раствор.
Осмотическое давление крови вычисляют криоскопическим методом с помощью
определения депрессии (точки замерзания), которая для крови составляет
0,56—0,58°С. Депрессия молярного раствора (раствор, в котором растворена 1
грамм-молекула вещества в 1
л воды) соответствует 1,86°С. Подставив значения в
уравнение Клапейрона, легко рассчитать, что осмотическое давление крови
равно приблизительно 7,6 атм.
Осмотическое давление крови зависит в основном от растворенных в ней
низкомолекулярных соединений, главным образом солей. Около 60% этого давления
создается NaCl. Осмотическое давление в крови, лимфе, тканевой жидкости, тканях
приблизительно одинаково и отличается постоянством. Даже в случаях, когда в
кровь поступает значительное количество воды или соли, осмотическое давление не
претерпевает существенных изменений. При избыточном поступлении в кровь
вода быстро выводится почками и переходит в ткани и клетки, что восстанавливает
исходную величину осмотического давления. Если же в крови повышается
концентрация солей, то в сосудистое русло переходит вода из тканевой жидкости, а
почки начинают усиленно выводить соли. Продукты переваривания белков, жиров и
углеводов, всасывающиеся в кровь и лимфу, а также низкомолекулярные продукты
клеточного метаболизма могут изменять осмотическое давление в небольших
пределах.
Поддержание постоянства осмотического давления играет чрезвычайно важную
роль в жизнедеятельности клеток.
Онкотическое давление. Является частью осмотического и зависит от
содержания крупномолекулярных соединений (белков) в растворе. Хотя концентрация
белков в плазме довольно велика, общее количество молекул из-за их большой
молекулярной массы относительно мало, благодаря чему онкотическое давление не
превышает 30
мм рт.ст. Онкотическое давление в большей степени зависит
от альбуминов (80% онкотического давления создают альбумины), что связано с
их относительно малой молекулярной массой и большим количеством молекул в
плазме.
Онкотическое давление играет важную роль в регуляции водного обмена. Чем
больше его величина, тем больше воды удерживается в сосудистом русле и тем
меньше ее переходит в ткани и наоборот. Онкотическое давление влияет на
образование тканевой жидкости, лимфы, мочи и всасывание воды в кишечнике.
Поэтому кровезамещающие растворы должны содержать в своем составе коллоидные
вещества, способные удерживать воду.
При снижении концентрации белка в плазме развиваются отеки, так как вода
перестает удерживаться в сосудистом русле и переходит в ткани.
Температура крови. Во многом зависит от интенсивности обмена веществ
того органа, от которого оттекает кровь, и колеблется в пределах 37—40°С. При
движении крови не только происходит некоторое выравнивание температуры в
различных сосудах, но и создаются условия для отдачи или сохранения тепла в
организме.
Концентрация водородных ионов и регуляция рН крови. В норме рН
крови соответствует 7,36, т. е. реакция слабоосновная. Колебания величины рН
крови крайне незначительны. Так, в условиях покоя рН артериальной крови
соответствует 7,4, а венозной — 7,34. В клетках и тканях рН достигает 7,2 и
даже 7,0, что зависит от образования в них в процессе обмена веществ «кислых»
продуктов метаболизма. При различных физиологических состояниях рН крови может
изменяться как в кислую (до 7,3), так и в щелочную (до 7,5) сторону. Более
значительные отклонения рН сопровождаются тяжелейшими последствиями для
организма. Так, при рН крови 6,95 наступает потеря сознания, и если эти
сдвиги в кратчайший срок не ликвидируются, то неминуема смерть. Если же
концентрация ионов Н+ уменьшается и рН становится равным 7,7, то
наступают тяжелейшие судороги (тетания), что также может привести к смерти.
В процессе обмена веществ ткани выделяют в тканевую жидкость, а
следовательно, и в кровь «кислые» продукты обмена, что должно приводить к сдвигу
рН в кислую сторону. Так, в результате интенсивной мышечной деятельности в
кровь человека может поступать в течение нескольких минут до
90 г
молочной кислоты. Если это количество молочной кислоты прибавить к объему
дистиллированной воды, равному объему циркулирующей крови, то концентрация ионов
Н+ возросла в ней в 40 000 раз. Реакция же крови при этих условиях
практически не изменяется, что объясняется наличием буферных систем крови. Кроме
того, в организме постоянство рН сохраняется за счет работы почек и легких,
удаляющих из крови СО2, избыток солей, кислот и оснований
(щелочей).
Постоянство рН крови поддерживается буферными системами: гемоглобиновой,
карбонатной, фосфатной и белками плазмы.
Самой мощной является буферная система гемоглобина. На ее долю
приходится 75% буферной емкости крови. Эта система включает восстановленный
гемоглобин (ННb) и калиевую соль восстановленного гемоглобина (КНb).
Буферные свойства системы обусловлены тем, что КНb как соль слабой кислоты
отдает ион К+ и присоединяет при этом ион Н+, образуя
слабодиссоциированную кислоту:
H+ + KHb =
K+ + HHb
Величина рН крови, притекающей к тканям, благодаря восстановленному
гемоглобину, способному связывать СО2 и Н+-ионы, остается
постоянной. В этих условиях ННЬ выполняет функции основания. В легких гемоглобин
ведет себя как кислота (оксигемоглобин ННbО2 является более сильной
кислотой, чем СО2), что предотвращает защелачивание крови.
Карбонатная буферная система
(H2CO3/NaHCO3) по своей мощности занимает
второе место. Ее функции осуществляются следующим образом: NaHCO3
диссоциирует на ионы Na+ и НСОз-. Если в кровь поступает
кислота более сильная, чем угольная, то происходит обмен ионами
Na+ с образованием слабодиссоциированной и легко растворимой угольной
кислоты, что предотвращает повышение концентрации ионов Н+ в
крови. Увеличение же концентрации угольной кислоты приводит к ее распаду (это
происходит под влиянием фермента карбоангидразы, находящегося в эритроцитах) на
Н2О и СО2. Последний поступает в легкие и выделяется в
окружающую среду. Если в кровь поступает основание, то она реагирует с
угольной кислотой, образуя натрия гидрокарбонат (NaНСОз) и воду, что опять-таки
препятствует сдвигу рН в щелочную сторону.
Фосфатная буферная система образована натрия дигидрофосфатом
(NaH2PO4) и натрия гидрофосфатом
(Na2HPO4). Первое соединение ведет себя как слабая
кислота, второе — как соль слабой кислоты. Если в кровь попадает более сильная
кислота, то она реагирует с Na2HPO4, образуя нейтральную
соль, и увеличивает количество слабодиссоциируемого
H++NaHPO4-=Na+
+
H2PO4-
Избыточное количество натрия дигидрофосфата при этом будет удаляться с мочой,
благодаря чему соотношение
NaH2PO4/Na2HPO4 не изменится.
Белки плазмы крови играют роль буфера, так как обладают амфотерными
свойствами: в кислой среде ведут себя как основания, а в основной — как
кислоты.
Важная роль в поддержании постоянства рН крови отводится нервной регуляции.
При этом преимущественно раздражаются хеморецепторы сосудистых рефлексогенных
зон, импульсы от которых поступают в продолговатый мозг и другие отделы ЦНС, что
рефлекторно включает в реакцию периферические органы — почки, легкие, потовые
железы, желудочно-кишечный тракт и др., деятельность которых направлена на
восстановление исходной величины рН. Так, при сдвиге рН в кислую сторону почки
усиленно выделяют с мочой анион Н2РО4- При
сдвиге рН крови в щелочную сторону увеличивается выделение почками анионов
НРО2- и НСОз-. Потовые железы человека способны выводить
избыток молочной кислоты, а легкие — СО2.
Буферные системы крови более устойчивы к действию кислот, чем оснований.
Основные соли слабых кислот, содержащиеся в крови, образует так называемый
щелочной резерв крови. Его величина определяется по тому количеству
СО2, которое может быть связано 100 мл крови при напряжении
СО2, равному 40
мм рт. ст.
При различных патологических состояниях может наблюдаться сдвиг рН как в
кислую, так и в щелочную сторону. Первый из них носит название ацидоза, второй —
алкалоза.
Суспензионная устойчивость крови (скорость оседания эритроцитов —
СОЭ). Кровь представляет собой суспензию, или взвесь, так как форменные
элементы ее находятся в плазме во взвешенном состоянии. Взвесь эритроцитов в
плазме поддерживается гидрофильной природой их поверхности, а также тем,
что эритроциты (как и другие форменные элементы) несут отрицательный заряд,
благодаря чему отталкиваются друг от друга. Если отрицательный заряд
форменных элементов уменьшается, что может быть обусловлено адсорбцией таких
положительно заряженных белков, как фибриноген, γ-глобулины,
парапротеины и др., то снижается электростатический «распор» между
эритроцитами. При этом эритроциты, склеиваясь друг с другом, образуют так
называемые монетные столбики. Одновременно положительно заряженные белки
выполняют роль межэритроцитарных мостиков. Такие «монетные столбики»,
застревая в капиллярах, препятствуют нормальному кровоснабжению тканей
и органов.
Если кровь поместить в пробирку, предварительно добавив в нее вещества,
препятствующие свертыванию, то через некоторое время можно увидеть, что кровь
разделилась на два слоя: верхний состоит из плазмы, а нижний представляет собой
форменные элементы, главным образом эритроциты. Исходя из этих свойств, Фарреус
предложил изучать суспензионную устойчивость эритроцитов, определяя
скорость их оседания в крови, свертываемость которой устранялась предварительным
добавлением цитрата натрия. Этот показатель получил наименование «скорость
оседания эритроцитов (СОЭ)».
Величина СОЭ зависит от возраста и пола. У новорожденных СОЭ равна 1—2 мм/ч,
у детей старше 1 года и у мужчин — 6—12 мм/ч, у женщин — 8—15 мм/ч, у пожилых
людей обоего пола — 15—20 мм/ч. Наибольшее влияние на величину СОЭ
оказывает содержание фибриногена: при увеличении его концентрации более 4
г/л СОЭ повышается. СОЭ резко увеличивается во время беременности, когда
содержание фибриногена в плазме значительно возрастает. Повышение СОЭ
наблюдается при воспалительных, инфекционных и онкологических заболеваниях,
а также при значительном уменьшении числа эритроцитов (анемия). Уменьшение
СОЭ у взрослых людей и детей старше 1 года является неблагоприятным
признаком.
Величина СОЭ зависит в большей степени от свойств плазмы, чем эритроцитов.
Так, если эритроциты мужчины с нормальной СОЭ поместить в плазму беременной
женщины, то эритроциты мужчины оседают с такой же скоростью, как и у женщин при
беременности.
6.2. ФОРМЕННЫЕ ЭЛЕМЕНТЫ КРОВИ
Все форменные элементы крови — эритроциты, лейкоциты и тромбоциты —
образуются в костном мозге из единой полипотентной, или плюрипотентной,
стволовой клетки (ПСК).
В костном мозге все кроветворные клетки собраны в грозди, которые окружены
фибробластами и эндотелиальными клетками. Созревшие клетки пробивают себе путь
среди расщелин, образованных фибробластами и эндотелием, в синусы, откуда
поступают затем в венозную кровь.
Несмотря на то, что все клетки крови являются потомками единой кроветворной
клетки, они несут различные специфические функции, в то же время общность
происхождения наделила их и общими свойствами. Так, все клетки крови, независимо
от их специфики, участвуют в транспорте различных веществ, выполняют защитные и
регуляторные функции.
6.2.1. Эритроциты
Эритроциты, или красные кровяные диски, впервые обнаружил в крови
лягушки Мальпиги (1661), а Левенгук (1673) показал, что они также присутствуют в
крови человека и млекопитающих.
В крови человека эритроциты имеют преимущественно форму двояковогнутого
диска. Поверхность диска в 1,7 раза больше, чем поверхность тела такого же
объема, но сферической формы; при этом диск умеренно изменяется без растяжения
мембраны клетки. Несомненно, форма двояковогнутого диска, увеличивая поверхность
эритроцита, обеспечивает транспорт большего количества различных веществ. Кроме
того, такая форма позволяет эритроцитам закрепляться в фибриновой сети при
образовании тромба. Но главное преимущество заключается в том, что форма
двояковогнутого диска обеспечивает прохождение эритроцита через капилляры. При
этом эритроцит перекручивается в узкой средней части, его содержимое из более
широкого конца перетекает к центру, благодаря чему эритроцит свободно входит в
узкий капилляр. Форма эритроцитов здоровых людей весьма вариабельна — от
двояковогнутой линзы до тутовой ягоды.
Эритроцит окружен плазматической мембраной, структура которой мало
отличается от таковой других клеток. Наряду с тем, что мембрана эритроцита
проницаема для катионов Na+ и К+, она особенно хорошо
пропускает 02, СО2, Сl- и
HCO3-. Цитоскелет в виде проходящих через клетку трубочек
и микрофиламентов в эритроците отсутствует, что придает ему эластичность и
деформируемость — столь необходимые свойства при прохождении через узкие
капилляры.
Размеры эритроцита весьма изменчивы, но в большинстве случаев их диаметр
равен 7,5—8,3 мкм, толщина — 2,1 мкм, площадь поверхности — 145 мкм2,
объем — 86 мкм3.
В норме число эритроцитов у мужчин равно 4—5*1012/л,
или 4 000 000—5 000 000 в 1 мкл. У женщин число эритроцитов меньше и, как
правило, не превышает 4,5*1012/л. При беременности число эритроцитов
может снижаться до 3,5-1012/л и даже до 3,0*1012 /л, и это
многие исследователи считают нормой.
У человека с массой тела 60
кг общее число эритроцитов равняется 25 триллионам. Если
положить все эритроциты одного человека один на другой, то получится «столбик»
высотой более 60
км.
В норме число эритроцитов подвержено незначительным колебаниям. При
различных заболеваниях количество эритроцитов может уменьшаться. Подобное
состояние носит название «эритропения» и часто сопутствует малокровию или
анемии. Увеличение числа эритроцитов обозначается как «эритроцитоз».
6.2.1.1. Гемоглобин и его
соединения
Основные функции эритроцитов обусловлены наличием в их составе особого белка
хромопротеида — гемоглобина. Молекулярная масса гемоглобина человека равна
68 800. Гемоглобин состоит из белковой (глобин) и железосодержащей (гем) частей.
На 1 молекулу глобина приходится 4 молекулы гема.
В крови здорового человека содержание гемоглобина составляет 120—165 г/л
(120—150 г/л для женщин и 130—160 г/л для мужчин). У беременных содержание
гемоглобина может понижаться до 110 г/л, что не является патологией.
Основное назначение гемоглобина — транспорт О2 и СО2.
Кроме того, гемоглобин обладает буферными свойствами, а также способностью
связывать некоторые токсичные вещества.
Гемоглобин человека и различных животных имеет разное строение. Это
касается белковой части — глобина, так как гем у всех представителей животного
мира имеет одну и ту же структуру. Гем состоит из молекулы порфирина, в центре
которой расположен ион Fe2+, способный присоединять О2.
Структура белковой части гемоглобина человека неоднородна, благодаря чему
белковая часть разделяется на ряд фракций. Большая часть гемоглобина
взрослого человека (95—98%) состоит из фракции А (от лат. adultus — взрослый);
от 2 до 3% всего гемоглобина приходится на фракцию А2; наконец, в
эритроцитах взрослого человека находится так называемый фетальный
гемоглобин (от лат. fetus — плод), или гемоглобин F, содержание которого в норме
подвержено значительным колебаниям, хотя редко превышает 1—2%. Гемоглобины А и
А2 обнаруживаются практически во всех эритроцитах, тогда как
гемоглобин F присутствует в них не всегда.
Гемоглобин F содержится преимущественно у плода. К моменту рождения ребенка
на его долю приходится 70—90%. Гемоглобин F имеет большее сродство к
О2, чем гемоглобин А, что позволяет тканям плода не испытывать
гипоксии, несмотря на относительно низкое напряжение О2 в его крови.
Эта приспособительная реакция объясняется тем, что гемоглобин F труднее вступает
в связь с 2,3-дифосфоглицернновой кислотой, которая уменьшает способность
гемоглобина переходить в оксигемоглобин, а следовательно, и обеспечивать
легкую отдачу О2 тканям.
Гемоглобин обладает способностью образовывать соединения с О2,
СО2 и СО. Гемоглобин, присоединивший О2, носит
наименование оксигемоглобина (ННbО2); гемоглобин, отдавший О2,
называется восстановленным, или редуцированным (ННb). В
артериальной крови преобладает содержание оксигемоглобина, от чего ее цвет
приобретает алую окраску. В венозной крови до 35% всего гемоглобина
приходится на ННb. Кроме того, часть гемоглобина через аминную группу
связывается с СО2, образуя карбогемоглобин
(ННbСО2), благодаря чему переносится от 10 до 20% всего
транспортируемого кровью СО2.
Гемоглобин способен образовывать довольно прочную связь с СО. Это соединение
называется карбоксигемоглобином (ННЬСО). Сродство гемоглобина к СО
значительно выше, чем к О2, поэтому гемоглобин, присоединивший СО,
неспособен связываться с О2. Однако при вдыхании чистого
О2 резко возрастает скорость распада карбоксигемоглобина, чем
пользуются на практике для лечения отравлений СО.
Сильные окислители (ферроцианид, бертолетова соль, пероксид, или перекись,
водорода и др.) изменяют заряд от Fe2+ до Fe3+, в
результате чего возникает окисленный гемоглобин — прочное соединение
гемоглобина с О2, носящее наименование метгемоглобина. При
этом нарушается транспорт О2, что приводит к тяжелейшим последствиям
для человека и даже смерти.
6.2.1.2. Цветовой показатель
О содержании в эритроцитах гемоглобина судят по так называемому
цветовому показателю, или фарб-индексу (Fi, от farb — цвет, index — показатель)
— относительной величине, характеризующей насыщение в среднем одного
эритроцита гемоглобином. Fi — процентное соотношение гемоглобина и эритроцитов,
при этом за 100% (или единиц) гемоглобина условно принимают величину, равную
166,7 г/л, а за 100% эритроцитов — 5*10 /л. Если у человека содержание
гемоглобина и эритроцитов равно 100%, то цветовой показатель равен 1. В норме Fi
колеблется в пределах 0,75—1,0 и очень редко может достигать 1,1. В этом случае
эритроциты называются нормохромными. Если Fi менее 0,7, то такие эритроциты
недонасыщены гемоглобином и называются гипохромными. При Fi более 1,1 эритроциты
именуются гиперхромными. В этом случае» объем эритроцита значительно
увеличивается, что позволяет ему содержать большую концентрацию гемоглобина. В
результате создается ложное впечатление, будто эритроциты перенасыщены
гемоглобином. Гипо- и гиперхромия встречаются лишь при анемиях. Определение
цветового показателя важно для клинической практики, так как позволяет
провести дифференциальный диагноз при анемиях различной этиологии.
6.2.1.3. Гемолиз
Гемолизом называется разрыв оболочки эритроцитов и выход гемоглобина в
плазму, благодаря чему кровь приобретает лаковый цвет. В искусственных условиях
гемолиз эритроцитов может быть вызван помещением их в гипотонический раствор.
Для здоровых людей минимальная граница осмотической стойкости соответствует
раствору, содержащему 0,42—0,48% NaCl, полный же гемолиз (максимальная
граница стойкости) происходит при концентрации 0,30— 0,34% NaCl. При анемиях
границы минимальной и максимальной стойкости смещаются в сторону повышения
концентрации гипотонического раствора.
Причины гемолиза. Гемолиз может быть вызван химическими агентами
(хлороформ, эфир, сапонин и др.), разрушающими мембрану эритроцитов. В
клинике нередко встречается гемолиз при отравлении уксусной кислотой.
Гемолизирующими свойствами обладают яды некоторых змей (биологический
гемолиз).
При сильном встряхивании ампулы с кровью также наблюдается разрушение
мембраны эритроцитов — механический гемолиз. Он может проявляться у
больных с протезированием клапанного аппарата сердца и сосудов. Кроме того,
механический гемолиз иногда возникает при длительной ходьбе (маршевая
гемоглобинурия) из-за травмирования эритроцитов в капиллярах стоп.
Если эритроциты заморозить, а потом отогреть, то возникает гемолиз,
получивший наименование термического. Наконец, при переливании
несовместимой крови и наличии аутоантител к эритроцитам развивается
иммунный гемолиз. Последний является причиной возникновения анемий и
нередко сопровождается выделением гемоглобина и его производных с мочой
(гемоглобинурия).
6.2.1.4. Функции
эритроцитов
Эритроцитам присуши три основные функции: транспортная, защитная и
регуляторная.
Транспортная функция эритроцитов заключается в том, что они
транспортируют О2 и CО2, аминокислоты, полипептиды, белки, углеводы, ферменты,
гормоны, жиры, холестерин, различные биологически активные соединения
(простагландины, лейкотриены и др.), микроэлементы и др.
Защитная функция эритроцитов заключается в том, что они играют
существенную роль в специфическом и неспецифическом иммунитете и принимают
участие в сосудисто-тромбоцитарном гемостазе, свертывании крови и
фибринолизе.
Регуляторную функцию эритроциты осуществляют благодаря
содержащемуся в них гемоглобину; регулируют рН крови, ионный состав плазмы
и водный обмен. Проникая в артериальный конец капилляра, эритроцит отдает воду и
растворенный в ней О2 и уменьшается в объеме, а переходя в
венозный конец капилляра, забирает воду, СО2 и продукты обмена,
поступающие из тканей и увеличивается в объеме.
Благодаря эритроцитам во многом сохраняется относительное постоянство состава
плазмы. Это касается не только солей. В случае увеличения концентрации в плазме
белков эритроциты их активно адсорбируют. Если же содержание белков в крови
уменьшается, то эритроциты отдают их в плазму.
Эритроциты являются носителями глюкозы и гепарина, обладающего
выраженным противосвертывающим действием. Эти соединения при увеличении их
концентрации в крови проникают через мембрану внутрь эритроцита, а при снижении
— вновь поступают в плазму.
Эритроциты являются регуляторами эритропоэза, так как в их составе содержатся
эритропоэтические факторы, поступающие при разрушении эритроцитов в костный мозг
и способствующие образованию эритроцитов. В случае разрушения эритроцитов
из освобождающегося гемоглобина образуется билирубин, являющийся одной
из составных частей желчи.
6.2.1.5. Эритрон. Регуляция
эритропоэза
Понятие «эритрон» введено английским терапевтом Каслом для обозначения массы
эритроцитов, находящихся в циркулирующей крови, в кровяных депо и костном мозге.
Принципиальная разница между эритроном и другими тканями организма заключается в
том, что разрушение эритроцитов осуществляется преимущественно макрофагами
за счет процесса, получившего наименование «эритрофагоцитоз». Образующиеся при
этом продукты разрушения и в первую очередь железо используются на построение
новых клеток. Таким образом, эритрон является замкнутой системой, в которой в
условиях нормы количество разрушающихся эритроцитов соответствует числу вновь
образовавшихся.
Развитие эритроцитов происходит в замкнутых капиллярах красного костного
мозга. Как только эритроцит достигает стадии ретикулоцита, он растягивает стенку
капилляра, благодаря чему сосуд раскрывается и ретикулоцит вымывается в
кровоток, где и превращается за 35—45 ч в молодой эритроцит — нормоцит. В норме
в крови содержится не более 1—2% ретикулоцитов.
В кровотоке эритроциты живут 80—120 дней. Продолжительность жизни эритроцитов
у мужчин несколько больше, чем у женщин.
Для нормального эритропоэза необходимо железо. Последнее поступает в
костный мозг при разрушении эритроцитов, из депо, а также с пищей и водой.
Взрослому человеку для нормального эритропоэза требуется в суточном рационе
12—15 мг железа. Железо откладывается в различных органах и тканях, главным
образом в печени и селезенке. Если железа в организм поступает недостаточно, то
развивается железодефицитная анемия.
Всасыванию железа в кишечнике способствует аскорбиновая кислота,
переводящая Fe3+ в Fe2+, который сохраняет
растворимость при нейтральных и щелочных значениях рН. На участке слизистой
оболочки тонкой кишки имеются рецепторы, облегчающие переход железа в энтероцит,
а оттуда в плазму. В слизистой оболочке тонкой кишки находится белок-переносчик
железа — трансферрин. Он доставляет железо в ткани, имеющие
трансферриновые рецепторы. В клетке комплекс трансферрина и железа распадается,
и железо вступает в связь с другим белком-переносчиком — ферритином.
Клетки-предшественники зрелых эритроцитов накапливают железо в ферритине. В
дальнейшем оно используется, когда клетка начинает образовывать большое
количество гемоглобина.
Важным компонентом эритропоэза является медь, которая усваивается
непосредственно в костном мозге и принимает участие в синтезе гемоглобина. Если
медь отсутствует, то эритроциты созревают лишь до стадии ретикулоцита. Медь
катализирует образование гемоглобина, способствуя включению железа в структуру
гема. Недостаток меди приводит к анемии.
Для нормального эритропоэза необходимы витамины и в первую
очередь витамин B12 и фолиевая кислота. Эти витамины оказывают
сходное взаимодополняющее действие на эритропоэз. Витамин B12
(внешний фактор кроветворения) синтезируется микроорганизмами, лучистыми
грибками и некоторыми водорослями. Для его образования необходим кобальт. В
организм человека витамин B12 поступает с пищей — особенно его много
в печени, мясе, яичном желтке.
Для всасывания витамина В12 требуется внутренний фактор
кроветворения, который носит наименование «гастромукопротеин». Это вещество
является комплексным соединением, образующимся в желудке.
Фолиевая кислота, или витамин В7, является водорастворимым
витамином, содержащимся во многих растительных продуктах, а также в печени,
почках, яйцах.
Витамин В12 и фолиевая кислота принимают участие в синтезе
глобина. Они обусловливают образование в эритробластах нуклеиновых кислот,
являющихся одним из основных строительных материалов клетки.
Немаловажную роль в регуляции эритропоэза играют другие витамины группы В, а
также железы внутренней секреции. Все гормоны, регулирующие обмен белков
(соматотропный гормон гипофиза, гормон щитовидной железы — тироксин и др.)
и кальция (паратгормон, тиреокальцитонин), необходимы для нормального
эритропоэза. Мужские половые гормоны (андрогены) стимулируют эритропоэз, тогда
как женские (эстрогены) — тормозят его, что обусловливает меньшее число
эритроцитов у женщин по сравнению с мужчинами.
Особо важную роль в регуляции эритропоэза играют специфические вещества,
получившие наименование «эритропоэтины». Еще в 1906 г. показано, что
сыворотка крови кроликов, перенесших кровопотерю, стимулирует электропоэз. В
дальнейшем было установлено, что эритропоэтины присутствуют в крови
животных и людей, испытывающих гипоксию — недостаточное поступление к тканям
кислорода, что наблюдается при анемиях, подъеме на высоту, мышечной работе,
снижении парциального давления кислорода в барокамере, заболеваниях сердца и
легких. В небольшой концентрации эритропоэтины обнаружены в крови здоровых
людей, что позволяет считать их физиологическими регуляторами эритропоэза.
Вместе с тем при анемиях, сопровождающих заболевания почек, эритропоэтины
отсутствуют или их концентрация значительно снижается. Эти данные позволили
предположить, что местом синтеза эритропоэтинов являются почки. Эритропоэтины
образуются также в печени, селезенке, костном мозге. Получены факты,
свидетельствующие о том, что мощной эритропоэтической активностью
обладают полипептиды эритроцитов, молекулярная масса которых не превышает
10 000.
Эритропоэтины оказывают действие непосредственно на клетки-предшественники
эритроидного ряда (КОЕ-Э – колониеобразующая единица эритроцитарная). Функции
эритропоэтинов сводятся к следующему: 1) ускорение и усиление перехода
стволовых клеток костного мозга в эритробласты; 2) увеличение числа митозов
клеток эритроидного ряда; 3) исключение одного или нескольких циклов
митотических делений; 4) ускорение созревания неделящихся клеток — нормобластов,
ретикулоцитов.
Ряд гемопоэтических факторов образуется стромой костного мозга и
костномозговыми фибробластами. «Микроокружение» костного мозга является
важнейшей частью кроветворного механизма. Эритроидные предшественники,
размещенные на ячеистой сети костномозговых фибробластов, быстро
развиваются и втискиваются между ними. Это объясняется тем, что для
дифференцировки эритроидных клеток требуется их плотное прикрепление (адгезия) к
окружающим структурам. Кроме того, фибробласты и эндотелиальные клетки являются
источником ростковых факторов кроветворения.
На эритропоэз действуют соединения, синтезируемые моноцитами,
макрофагами, лимфоцитами и другими клетками, получившие название
«интерлейкины». Согласно международной классификации, они
обозначаются арабскими цифрами (ИЛ-1, ИЛ-2 и т. д.). На полипотентную стволовую
клетку (ПСК) непосредственно влияют и способствуют ее дифференцировке ИЛ-3,
ИЛ-6, ИЛ-11 и ИЛ-12. В частности, активированные макрофаги выделяют ИЛ-1, а
также фактор некроза опухолей (ФНО). ИЛ-1 и ФНО стимулируют фибробласты и
эндотелиальные клетки, благодаря чему они усиленно продуцируют так называемый
белковый фактор Стала, оказывающий влияние непосредственно на ПСК и
способствующий ее дифференцировке. Кроме того, фибробласты, эндотелиальные
клетки и активированные Т-лимфоциты способны выделять ИЛ-6, ИЛ-11 и
гранулоцитарно-макрофагальный колониестимулирующий фактор (ГМ-КСФ). Фактор
Стала, ИЛ-3, ИЛ-6, ИЛ-11, ИЛ-1 и ГМ-КСФ относятся к раннедействующим
гемопоэтическим ростовым факторам. По мере того как родоначальники нескольких
линий кроветворных клеток дифференцируются в родоначальники одной линии, в
реакцию вступают позднедействующие гемопоэтические ростовые факторы и
эритропоэтин.
Важная роль в эритропоэзе принадлежит ядерным факторам — ГАТА-1
(внутриядерный регулятор транскрипции в эритроне) и НФЕ-2. Отсутствие ГАТА-1
предотвращает образование эритроцитов, недостаток НФЕ-2 нарушает всасывание
железа в кишечнике и синтез глобина.
6.2.2. Лейкоциты
Лейкоциты, или белые кровяные тельца, представляют собой образования
различной формы и величины. По строению лейкоциты делят на две большие группы:
зернистые, или гранулоциты, и незернистые, или
агранулоциты. К гранулоцитам относятся нейтрофилы, эозинофилы и базофилы,
к агранулоцитам — лимфоциты и моноциты. Свое наименование клетки зернистого ряда
получили от способности окрашиваться красками: эозинофилы воспринимают кислую
краску (эозин), базофилы — щелочную (гематоксилин), а нейтрофилы — и ту, и
другую.
В норме количество лейкоцитов у взрослых людей колеблется от 4,5 до 8,5 тыс.
в 1 мм3, или 4,5—8,5*109/л.
Увеличение числа лейкоцитов носит название лейкоцитоза, уменьшение —
лейкопении. Лейкоцитозы могут быть физиологические и патологические,
тогда как лейкопении встречаются только при патологии.
6.2.2.1. Физиологические
лейкоцитозы. Лейкопении
Различают следующие виды физиологических лейкоцитозов:
Пищевой. Возникает после приема пищи. При этом число
лейкоцитов увеличивается незначительно (в среднем на 1—3 тыс. в мкл) и
редко выходит за границу верхней физиологической нормы. При пищевом лейкоцитозе
большое количество лейкоцитов скапливается в подслизистой основе тонкой
кишки. Здесь они осуществляют защитную функцию — препятствуют попаданию
чужеродных агентов в кровь и лимфу. Пищевой лейкоцитоз носит
перераспределительный характер и обеспечивается поступлением лейкоцитов в
кровоток из депо крови.
Миогенный. Наблюдается после выполнения тяжелой мышечной
работы. Число лейкоцитов при этом может возрастать в 3—5 раз. Огромное
количество лейкоцитов при физической нагрузке скапливается в мышцах.
Миогенный лейкоцитоз носит как перераспределительный, так и истинный
характер, так как при нем наблюдается усиление костномозгового
кроветворения.
Эмоциональный. Как и лейкоцитоз при болевом раздражении, носит
перераспределительный характер и редко достигает высоких показателей.
При беременности. Большое количество лейкоцитов скапливается в
подслизистой основе матки. Этот лейкоцитоз в основном носит местный характер.
Его физиологический смысл состоит не только в предупреждении попадания инфекции
в организм роженицы, но и в стимулировании сократительной функции матки.
Лейкопении встречаются только при патологических состояниях. Особенно
тяжелая лейкопения может наблюдаться в случае поражения костного мозга — острых
лейкозах и лучевой болезни. При этом изменяется функциональная активность
лейкоцитов, что приводит к нарушениям в специфической и неспецифической
защите, попутным заболеваниям, часто инфекционного характера, и даже
смерти.
6.2.2.2. Лейкоцитарная формула
В норме и патологии учитывается не только количество лейкоцитов, но и их
процентное соотношение, получившее наименование лейкоцитарной формулы, или
лейкограммы (табл. 6.3).
Таблица 6.3. Лейкоцитарная формула здорового человека, %
|
Гранулоциты |
Агранулоциты |
|
нейтрофилы |
базофилы
|
эозинофилы
|
лимфоциты
|
моноциты
|
|
юные |
палочко-ядерные |
сегменто-ядерные |
|
0—1 |
1—4 |
45—65 |
0—1 |
1—4 |
25—40 |
2—8 |
В крови здорового человека могут встречаться зрелые и юные формы лейкоцитов,
однако в норме обнаружить их удается лишь у самой многочисленной группы —
нейтрофилов. К ним относятся юные и палочкоядерные нейтрофилы. Юные нейтрофилы,
или миелоциты, имеют довольно крупное бобовидное ядро, палочкоядерные —
содержат ядро, не разделенное на отдельные сегменты. Зрелые, или
сегментоядерные, нейтрофилы имеют ядро, разделенное на 2 или 3 сегмента. Чем
больше сегментов в ядре, тем старее нейтрофил. Увеличение количества юных и
палочкоядерных нейтрофилов свидетельствует об омоложении крови и носит
название сдвига лейкоцитарной формулы влево, снижение количества этих
клеток свидетельствует о старении крови и называется сдвигом лейкоцитарной
формулы вправо. Сдвиг влево часто наблюдается при лейкозах (белокровие),
инфекционных и воспалительных заболеваниях.
6.2.2.3. Характеристика
отдельных видов лейкоцитов
Нейтрофилы. Созревая в костном мозге, задерживаются в нем на 3—5
дней, составляя костномозговой резерв гранулоцитов.
Лейкопоэз осуществляется экстраваскулярно и лейкоциты, в том числе и
нейтрофилы, попадают в сосудистое русло благодаря амебовидному движению и
выделению протеолитических ферментов, способных растворять белки костного мозга
и капилляров. В циркулирующей крови нейтрофилы живут от 8 ч до 7 сут.
Находящиеся в кровотоке нейтрофилы могут быть условно разделены на 2 группы: 1)
свободно циркулирующие и 2) занимающие краевое положение в сосудах. Между обеими
группами существует динамическое равновесие и постоянный обмен.
Следовательно, в сосудистом русле нейтрофилов содержится приблизительно в 2 раза
больше, чем определяется в вытекающей крови.
Предполагают, что разрушение нейтрофилов происходит за пределами
сосудистого русла. По-видимому, все лейкоциты уходят в ткани, где и погибают.
Обладая фагоцитарной функцией, нейтрофилы поглощают бактерии и продукты
разрушения тканей. В составе нейтрофилов содержатся ферменты, разрушающие
бактерии. Нейтрофилы способны адсорбировать антитела и переносить их к
очагу воспаления, принимают участие в обеспечении иммунитета.
Под влиянием продуктов, выделяемых нейтрофилами, усиливается
митотическая активность клеток, ускоряются процессы репарации,
стимулируется гемопоэз и растворение фибринового сгустка.
Базофилы. В крови базофилов очень мало (40—60 в 1 мкл) однако в
различных тканях, в том числе сосудистой стенке, содержатся тучные клетки,
иначе называемые «тканевые базофилы». Функция базофилов обусловлена наличием в
них ряда биологически активных веществ. К ним в первую очередь принадлежит
гистамин, расширяющий кровеносные сосуды. В базофилах содержатся
противосвертывающее вещество гепарин, а также гиалуроновая кислота, влияющая на
проницаемость сосудистой стенки. Кроме того, базофилы содержат фактор
активации тромбоцитов — ФАТ (соединение, обладающее чрезвычайно широким спектром
действия), тромбоксаны (соединения, способствующие агрегации тромбоцитов),
лейкотриены и простагландины — производные арахидоновой кислоты и др. Особо
важную роль играют эти клетки при аллергических реакциях (бронхиальная
астма, крапивница, глистные инвазии, лекарственная болезнь и др.), когда
под влиянием комплекса антиген — антитело происходит дегрануляция базофилов и
биологически активные соединения поступают в кровь, обусловливая
клиническую картину перечисленных заболеваний.
Количество базофилов резко возрастает при лейкозах, стрессовых ситуациях и
слегка увеличивается при воспалении.
Эозинофилы. Длительность пребывания эозинофилов в кровотоке не
превышает нескольких часов, после чего они проникают в ткани, где и разрушаются.
Эозинофилы обладают фагоцитарной активно- стью. Особенно интенсивно они
фагоцитируют кокки. В тканях эозинофилы скапливаются преимущественно в тех
органах, где содержится гистамин — в слизистой оболочке и полслизистой
основе желудка и тонкой кишки, в легких. Эозинофилы захватывают гистамин и
разрушают его с помощью фермента гистаминазы. В составе эозинофилов
находится фактор, тормозящий выделение гистамина тучными клетками и базофилами.
Эозинофилы играют важную роль в разрушении токсинов белкового
происхождения, чужеродных белков и иммунных комплексов.
Чрезвычайно велика роль эозинофилов, осуществляющих цитотоксический эффект, в
борьбе с гельминтами, их яйцами и личинками. В частности, при контакте
активированного эозинофила с личинками происходит его дегрануляция с последующим
выделением большого количества белка и ферментов, например пероксидаз, на
поверхность личинки, что приводит к разрушению последней. Увеличение числа
эозинофилов, наблюдаемое при миграции личинок, является одним из важнейших
механизмов в ликвидации гельминтозов.
Содержание эозинофилов резко возрастает при аллергических заболеваниях, когда
происходит дегрануляция базофилов и выделение анафилактического
хемотаксического фактора, который привлекает эозинофилы. При этом
эозинофилы выполняют роль «чистильщиков», фагоцитируя и инактивируя
продукты, выделяемые базофилами.
В эозинофилах содержатся катионные белки, которые активируют компоненты
калликреин-кининовой системы и влияют на свертывание крови. Предполагают,
что катионные белки, повреждая эндотелий, играют важную роль при развитии
некоторых видов патологии сердца и сосудов.
При тяжело протекающих инфекционных заболеваниях число эозинофилов резко
снижается, а иногда при подсчете лейкоцитарной формулы они вообще не
выявляются (развивается анэозинопения).
Моноциты. Циркулируют до 70 ч, а затем мигрируют в ткани, где образуют
обширное семейство тканевых макрофагов. Функции их весьма многообразны. Моноциты
являются чрезвычайно активными фагоцитами, распознают антиген и переводят
его в так называемую иммуногенную форму, образуют биологически активные
соединения — монокины (действующие в основном на лимфоциты), играют существенную
роль в противоинфекционном и противораковом иммунитете, синтезируют
отдельные компоненты системы комплемента, а также факторы, принимающие участие в
сосудисто-тромбоцитарном гемостазе, процессе свертывания крови и
растворении кровяного сгустка.
Лимфоциты. Как и другие виды лейкоцитов, образуются в костном
мозге, а затем поступают в сосудистое русло. Здесь одна популяция лимфоцитов
направляется в вилочковую железу, где превращается в так называемые Т-лимфоциты
(от слова thymus).
Популяция Т-лимфоцитов гетерогенна и представлена следующими классами клеток.
Т-киллеры, или убийцы (от англ. tu kill — убивать), осуществляющие лизис
клеток-мишеней, к которым можно отнести возбудителей инфекционных болезней,
грибки, микобактерии, опухолевые клетки и др. Т-хелперы, или помощники
иммунитета. Различают Т—Т-хелперы, усиливающие клеточный
иммунитет, и Т—В-хелперы, облегчающие течение гуморального
иммунитета. Т-амплифайеры усиливают функцию Т- и В-лимфоцитов, однако в
большей степени влияют на Т-лимфоциты. Т-супрессоры — лимфоциты,
препятствующие иммунному ответу. Различают Т—Т-супрессоры, подавляющие
клеточный иммунитет, и Т—В-супрессоры, угнетающие гуморальный
иммунитет. Т-дифференцирующие, или Td-лимфоциты, регулируют функцию
стволовых кроветворных клеток, т. е. влияют на соотношение эритроцитарного,
лейкоцитарного и тромбоцитарного (мегакариоцитарного) ростков костного
мозга. Т-контрсупрессоры препятствуют действию Т-супрессоров и,
следовательно, усиливают иммунный ответ. Т-клетки памяти хранят
информацию о ранее действующих антигенах и таким образом регулируют так
называемый вторичный иммунный ответ, который проявляется в более короткие сроки,
так как минует основные стадии этого процесса.
Другая популяция лимфоцитов образует В-лимфоциты (от слова bursa),
окончательное формирование которых у человека и млекопитающих, по-видимому,
происходит в костном мозге или системе лимфоидно-эпителиальных образований,
расположенных по ходу тонкой кишки (лимфоидные, или пейеровы бляшки и
др.).
Большинство В-лимфоцитов в ответ на действие антигенов и цитокинов переходит
в плазматические клетки, вырабатывающие антитела и потому именуемые
антителопродуцентами. Среди В-лимфоцитов также различают В-киллеры, В-хелперы и
В-супрессоры.
В-киллеры выполняют те же функции, что и Т-киллеры. Что касается
В-хелперов, то они способны представлять антиген, усиливать действие
Td-лимфоцитов и Т-супрессоров, а также участвовать в других реакциях
клеточного и гуморального иммунитета. Функция В-cynpeccopoв
заключается в торможении пролиферации антителопродуцентов, к которым
принадлежит основная масса В-лимфоцитов.
Существует группа клеток, получивших наименование «ни Т-, ни В-лимфоциты». К
ним относятся так называемые 0-лимфоциты, являющиеся предшественниками Т- и
В-клеток и составляющие их резерв. Большинство исследователей относят к
0-лим-фоцитам особые клетки, именуемые натуральными (природными) киллерами, или
НК-лимфоцитами. Как и другие цитотоксические лимфоциты (ЦТЛ), НК-лимфоциты
секретируют белки, способные «пробуравливать» отверстия (поры) в мембране
чужеродных клеток и потому названные перфоринами. ЦТЛ содержат протеолитические
ферменты (цитолизины), которые проникают в чужеродную клетку через образующиеся
поры и разрушают ее.
Существуют клетки, несущие на своей поверхности маркеры Т- и В-лимфоцитов
(двойные клетки). Они способны заменять как те, так и другие.
6.2.2.4. Регуляция лейкопоэза
Все лейкоциты образуются в красном костном мозге из единой стволовой клетки,
однако родоначальницей миелопоэза является бипотенциальная колониеобразующая
единица гранулоцитарно-моноцитарная (КОЕ-ГМ) или клетка-предшественница. Для
ее роста и дифференцировки необходим особый колониестимулирующий фактор
(КСФ), вырабатываемый у человека моноцитарно-макрофагальными клетками,
костным мозгом и лимфоцитами.
КСФ является гликопротеидом и состоит из двух частей — стимулятора продукции
эозинофилов (Эо-КСФ) и стимулятора продукции нейтрофилов и моноцитов
(ГМ-КСФ), относящихся к ранним гемопоэтическим ростовым факторам. Содержание
ГМ-КСФ стимулируется Т-хелперами и подавляется Т-супрессорами. На более
поздних этапах на лейкопоэз влияют гранулоцитарный колониестимулирующий
фактор — Г-КСФ (способствует развитию нейтрофилов) и макрофагальный
колониестимулирующий фактор — М-КСФ (приводит к образованию моноцитов),
являющиеся позднодействующими специфическими ростовыми факторами.
Установлено, что Td-лимфоциты стимулируют дифференцировку клеток в
гранулоцитарном направлении. В регуляции размножения ранних поли- и унипотентных
клеток имеет важное значение их взаимодействие с Т-лимфоцитами и макрофагами.
Эти клетки влияют на клетки-предшественницы с помощью лимфокинов и
монокинов, содержащихся в мембране и отделяющихся от нее в виде «пузырьков» при
тесном контакте с клетками-мишенями.
Из костного мозга и отдельных видов лейкоцитов (гранулоцитов и агранулоцитов)
выделен комплекс полипептидных факторов, выполняющих функции специфических
лейкопоэтинов.
Важная роль в регуляции лейкопоэза отводится интерлейкинам. В частности, ИЛ-3
не только стимулирует гемопоэз, но и является фактором роста и развития
базофилов. ИЛ-5 необходим для роста и развития эозинофилов. Многие интерлейкины
(ИЛ-2, ИЛ-4, ИЛ-6, ИЛ-7 и др.) являются факторами роста и дифференцировки Т- и
В-лимфоцитов (см. раздел 6.2.2.8).
Лейкоциты являются наиболее «подвижной» частью крови, быстро реагирующей на
различные изменения в окружающей среде и организме развитием лейкоцитоза,
что обеспечивается существованием клеточного резерва. Известны два типа
гранулоцитарных резервов — сосудистый и костномозговой. Сосудистый
гранулоцитарный резерв представляет собой большое количество гранулоцитов,
расположенных вдоль стенок сосудистого русла, откуда они мобилизуются при
повышении тонуса симпатического отдела автономной (вегетативной) нервной
системы.
Количество клеток костномозгового гранулоцитарного резерва в 30—50 раз
превышает их количество в кровотоке. Мобилизация этого резерва происходит при
инфекционных заболеваниях, сопровождается сдвигом лейкоцитарной формулы
влево и обусловлена в основном воздействием эндотоксинов.
Своеобразные изменения претерпевают лейкоциты в разные стадии
адаптационного синдрома, что обусловлено действием гормонов гипофиза (АКТГ) и
надпочечника (адреналина, кортизона, дезоксигидрокортизона). Уже через несколько
часов после стрессорного воздействия развивается лейкоцитоз, который обусловлен
выбросом нейтрофилов, моноцитов и лимфоцитов из депо крови. При этом число
лейкоцитов не превышает 16—18 тыс. в 1 мкл. В стадии резистентности число и
состав лейкоцитов мало отличаются от нормы. В стадии истощения развивается
лейкоцитоз, сопровождающийся увеличением числа нейтрофилов и снижением
числа лимфоцитов и эозинофилов.
6.2.2.5. Неспецифическая
резистентность и иммунитет
Основное назначение лейкоцитов — участие в защитных реакциях организма
против чужеродных агентов, способных нанести ему вред. Различают специфическую
защиту, или иммунитет, и неспецифическую резистентность организма.
Последняя, в отличие от иммунитета, направлена на уничтожение любого чужеродного
агента. К неспецифической резистентности относятся фагоцитоз и пиноцитоз,
система комплемента, естественная цитотоксичность, действие интерферонов
лизоцима, β-лизинов и других гуморальных факторов защиты.
Фагоцитоз. Это поглощение чужеродных частиц или клеток и их дальнейшее
уничтожение. Явление фагоцитоза открыто И. И. Мечниковым, за что ему была
присуждена Нобелевская премия 1908 г. Фагоцитоз присущ нейтрофилам,
эозинофилам, моноцитам и макрофагам.
И. И. Мечников выделил следующие стадии фагоцитоза: 1) приближение
фагоцита к фагоцитируемому объекту, или лиганду; 2) контакт лиганда с мембраной
фагоцита; 3) поглощение лиганда; 4) переваривание или уничтожение
фагоцитированного объекта.
Всем фагоцитам присуща амебовидная подвижность. Сцепление с субстратом, к
которому движется лейкоцит, носит название адгезии. Только фиксированные,
или адгезированные, лейкоциты способны к фагоцитозу.
Фагоцит может улавливать отдаленные сигналы (хемотаксис) и мигрировать в их
направлении (хемокинез). Хотя сотни продуктов метаболизма влияют на подвижность
лейкоцитов, их действие проявляется лишь в присутствии особых соединений —
хемоаттрактантов. К хемоаттрактантам относят продукты распада соединительной
ткани, иммуноглобулинов, фрагменты активных компонентов комплемента,
некоторые факторы свертывания крови и фибринолиза, простагландины, лейкотриены,
лимфокины и монокины. Благодаря хемотаксису, фагоцит целенаправленно движется в
сторону повреждающего агента. Чем выше концентрация хемоаттрактанта, тем большее
число фагоцитов устремляется в зону повреждения и тем с большей скоростью они
движутся. Для взаимодействия с хемоаттрактантом у фагоцита имеются специфические
гликопротеиновые образования — рецепторы; их число на одном нейтрофиле достигает
2 • 103—2 • 105. Движение фагоцитов осуществляется в результате
взаимодействия актина и миозина и сопровождается выдвижением
псевдоподий, которые служат точкой опоры при перемещении фагоцита.
Прикрепляясь к субстрату, псевдоподия перетягивает фагоцит на новое место.
Двигаясь таким образом, лейкоцит проходит через эндотелий капилляра; прилипая
к сосудистой стенке, он выпускает псевдоподию, которая пронизывает стенку
сосуда. В этот выступ постепенно «переливается» тело лейкоцита. После этого
лейкоцит отделяется от стенки сосуда и может передвигаться в тканях.
Контакт фагоцита с фагоцитируемым объектом может быть обусловлен
разностью электрических зарядов, повышенной степенью гидрофобности или
гидрофильностью лиганда, наличием на его поверхности лектинов, способных
специфически связываться с мембранной манозой или инсулином макрофага. В
большинстве случаев контакт опосредуется особыми соединениями — опсонинами,
значительно усиливающими фагоцитоз. К последним относятся иммунные
комплексы, некоторые фрагменты системы комплемента (см. раздел 6.2.2.6),
С-реактивный белок, агрегированные белки, фибронектины и др. Наиболее детально
опосредованный фагоцитоз изучен с участием гликопротеина фибронектина
(молекулярная масса 440 000), обладающего значительной клейкостью, что облегчает
взаимодействие фагоцита и лиганда. Фибронектин находится в
нерастворимой форме в соединительной ткани и в растворимой — в
α2-глобулиновой фракции плазмы. Кроме того, во взаимодействии
фагоцита и фагоцитируемого объекта принимают участие близкий по строению к-
фибронектину белок ламинин, а также ионы Са2+ и Mg2+ . Эта реакция
обеспечивается наличием на мембране фагоцитов специфических рецепторов.
Как только лиганд взаимодействует с рецептором, наступает конформация
последнего и сигнал передается на фермент, связанный с рецептором в единый
комплекс, благодаря чему осуществляется поглощение фагоцитируемого объекта.
Существует несколько механизмов поглощения, но все они сводятся к тому,
что лиганд оказывается заключенным в мембрану фагоцита. Образующаяся при этом
фагосома передвигается к центру клетки, где сливается с лизосомами, в результате
чего появляется фаголизосома. В последней, фагоцитируемый объект может
погибнуть. Это так называемый завершенный фагоцитоз. Но нередко
встречается незавершенный фагоцитоз, когда фагоцитируемый объект
может жить и развиваться в фагоците. Подобное явление наблюдается при
некоторых инфекционных заболеваниях — туберкулезе, гонорее, менингококковой
и вирусной инфекциях.
Последняя стадия фагоцитоза — уничтожение лиганда. Основным «оружием»
фагоцитов являются продукты частичного восстановления кислорода — пероксид
водорода, и свободные радикалы. Они вызывают пероксидное окисление липидов,
белков и нуклеиновых кислот, благодаря чему повреждается мембрана клетки.
В момент контакта рецепторов с фагоцитируемым объектом наступает
активация оксидаз — мембранных ферментов, переносящих электроны на кислород и
отнимающих их у восстановленных молекул. При образовании фаголизосомы
происходит резкое усиление окислительных процессов внутри нее, в результате чего
наступает гибель бактерий.
В процессе фагоцитоза утилизируемый клетками кислород превращается в
супероксидный анион-радикал (О2- ). В результате окисления
НАДФ*Н2 усиленно генерируется пероксид водорода, которому
присуще сильное окислительное действие. Фагоциты обладают универсальным
свойством высвобождать супероксидные радикалы, прежде всего
О2-.
На фагоцитируемый объект, заключенный в фагосому или фаголизосому, по системе
микротрубочек изливаются содержимое гранул, а также образовавшиеся
метаболиты. В частности, миелопероксидаза нейтрофилов, окисляя мембранные белки,
способна инактивировать грамположительные и грамотрицательные бактерии, вирусы,
грибки, микоплазмы при обязательном участии галогенов (анионов Сl- и
пероксида водорода (Н2О2). В уничтожении бактерий внутри
фагоцита принимает участие фермент лизоцим (мурамидаза), вызывающий гидролиз
гликопротеидов оболочки. В гранулоцитах содержится уникальная субстанция —
фагоцитин, обладающая антибактериальным действием и способная уничтожить
грамотрицательную и грамположительную микрофлору.
К другим механизмам, приводящим к гибели фагоцитируемого объекта, относятся
действие катионных белков, меняющих поверхностные свойства мембраны;
влияние лактоферрина, конкурирующего за ионы железа; действие различных
амилолитических, протеолитических и липолитических ферментов, содержащихся в
гранулах фагоцитов и разрушающих мембрану бактерий и вирусов.
Система комплемента. Комплемент — ферментная система, состоящая
более чем из 20 белков, играющая важную роль в осуществлении защитных
реакций, течении воспаления и разрушения (лизиса) мембран бактерий и различных
клеток.
При активации системы комплемента усиливается разрушение чужеродных и старых
клеток, активируются фагоцитоз и течение иммунных реакций, повышается
проницаемость сосудистой стенки, ускоряется свертывание крови, что в конечном
итоге приводит к более быстрой ликвидации патологического процесса.
Иммунитет. Это комплекс реакций, направленных на поддержание
гомеостаза при встрече организма с агентами, которые расцениваются как
чужеродные независимо от того, образуются ли они в самом организме или поступают
в него извне.
Чужеродные для данного организма соединения, способные вызывать иммунный
ответ, получили наименование «антигены» (АГ). Теоретически любая молекула может
быть АГ. В результате действия АГ в организме образуются антитела (AT),
сенсибилизируются (активируются) лимфоциты, благодаря чему они
приобретают способность принимать участие в иммунном ответе.
Специфичность АГ заключается в том, что он избирательно реагирует с
определенными AT или лимфоцитами, появляющимися после попадания АГ в
организм.
Способность АГ вызывать специфический иммунный ответ обусловлена
наличием на его молекуле многочисленных детерминант (эпитопов), к которым
специфически, как ключ к замку, подходят активные центры (антидетерминанты)
образующихся AT. АГ, взаимодействуя со своими AT, образуют иммунные
комплексы (ИК). Как правило, АГ — это молекулы с высокой молекулярной массой;
существуют потенциально активные в иммунологическом отношении вещества, величина
молекулы которых соответствует одной отдельной антигенной детерминанте.
Такие молекулы носят наименование гаптенов. Последние способны вызывать
иммунный ответ, только соединяясь с полным АГ, т. е. белком.
Органы, принимающие участие в иммунитете, делят на четыре группы.
1. Центральные — тимус, или вилочковая железа, и, по-видимому, костный
мозг.
2. Периферические, или вторичные, — лимфатические узлы, селезенка, система
лимфоэпителиальных образований, расположенных в слизистых оболочках
различных органов.
3. Забарьерные — ЦНС, семенники, глаза, паренхима тимуса и при беременности —
плод.
4. Внутрибарьерные — кожа.
Различают клеточный и гуморальный иммунитет. Клеточный иммунитет направлен на
уничтожение чужеродных клеток и тканей и обусловлен действием Т-киллеров.
Типичным примером клеточного иммунитета является реакция отторжения чужеродных
органов и тканей, в частности кожи, пересаженной от человека человеку.
Гуморальный иммунитет обеспечивается образованием AT и обусловлен в основном
функцией В-лимфоцитов.
Иммунный ответ. В иммунном ответе принимают участие иммунокомпетентные
клетки, которые могут быть разделены на антигенпрезентирующие (представляющие
АГ), регуляторные (регулирующие течение иммунных реакций) и эффекторы
иммунного ответа (осуществляющие заключительный этап в борьбе с АГ).
К антигенпрезентирующим клеткам относятся моноциты и макрофаги,
эндотелиальные клетки, пигментные клетки кожи (клетки Лангерганса) и др. К
регуляторным клеткам относятся Т- и В-хелперы, супрессоры, контрсупрессоры,
Т-лимфоциты памяти. Наконец, к эффекторам иммунного ответа принадлежат
Т- и В-киллеры и В-лимфоциты, являющиеся в основном антителопродуцентами.
Важная роль в иммунном ответе отводится особым цитокинам, получившим
наименование интерлейкинов (ИЛ). Из названия видно, что ИЛ обеспечивает
взаимосвязь отдельных видов лейкоцитов в иммунном ответе. Они представляют собой
малые белковые молекулы с молекулярной массой 15 000—30 000.
ИЛ-1 — соединение, выделяемое при антигенной стимуляции моноцитами,
макрофагами и другими антигенпрезентирующими клетками. Его действие в основном
направлено на Т-хелперы (амплифайеры) и макрофаги-эффекторы. ИЛ-1 стимулирует
гепатоциты, благодаря чему в крови возрастает концентрация белков,
получивших наименование реактантов острой фазы, так как их
содержание всегда увеличивается в острую фазу воспаления. К таким белкам
относятся фибриноген, С-реактивный белок, α1-антитрипсин и др. Белки острой фазы
воспаления играют важную роль в репарации тканей, связывают
протеолитические ферменты, регулируют клеточный и гуморальный иммунитет.
Увеличение концентрации реактантов острой фазы является приспособительной
реакцией, направленной на ликвидацию патологического процесса. Кроме того,
ИЛ-1 усиливает фагоцитоз, а также ускоряет рост кровеносных сосудов в зонах
повреждения.
ИЛ-2 выделяется Т-амплифайерами под воздействием ИЛ-1 и АГ; является
стимулятором роста для всех видов Т-лимфоцитов (киллеров, хелперов, супрессоров)
и активатором НК-клеток.
ИЛ-3 выделяется стимулированными Т-хелперами, моноцитами и
макрофагами. Его действие направлено преимущественно на рост и развитие тучных
клеток и базофилов, а также предшественников Т- и В-лимфоцитов.
ИЛ-4 продуцируется в основном стимулированными Т-хелперами и обладает
чрезвычайно широким спектром действия, так как способствует росту и
дифференцировке В-лимфоцитов, активирует макрофаги, Т-лимфоциты и тучные
клетки, индуцирует продукцию иммуноглобулинов отдельных классов.
ИЛ-5 выделяется стимулированными Т-хелперами и является фактором
пролиферации и дифференцировки эозинофилов, а также В-лимфоцитов.
ИЛ-6 продуцируется стимулированными моноцитами, макрофагами,
эндотелием, Т-хелперами и фибробластами; вместе с ИЛ-4 обеспечивает рост и
дифференцировку В-лимфоцитов, способствуя их переходу в антителопродуценты, т.
е. плазматические клетки.
ИЛ-7 первоначально выделен из стромальных клеток костного мозга;
усиливает рост и пролиферацию Т- и В-лимфоцитов, а также влияет на развитие
тимоцитов в тимусе.
ИЛ-8 образуется стимулированными моноцитами и макрофагами. Его
назначение сводится к усилению хемотаксиса и фагоцитарной активности
нейтрофилов.
ИЛ-9 продуцируется Т-лимфоцитами и тучными клетками. Действие его
направлено на усиление роста Т-лимфоцитов. Кроме того, он способствует развитию
эритроидных колоний в костном мозге.
ИЛ-10 образуется макрофагами и усиливает пролиферацию зрелых и
незрелых тимоцитов, а также способствует дифференцировке Т-киллеров.
ИЛ-11 продуцируется стромальными клетками костного мозга. Играет
важную роль в гемопоэзе, особенно тромбоцитопоэзе.
ИЛ-12 усиливает цитотоксичность Т-киллеров и НК-лимфоцитов.
Иммунный ответ начинается с взаимодействия антигеипрезентирующих клеток с АГ,
после чего происходят его фагоцитоз и переработка до продуктов деградации,
которые выделяются наружу и оказываются за пределами антигенпрезентирующей
клетки.
Специфичность иммунного ответа обеспечивается наличием особых антигенов,
получивших у мышей наименование la-белка. У человека его роль выполняют
человеческие лейкоцитарные антигены II класса, тип DR (Human Leukocyte Antigens,
или HLA).
la-белок находится практически на всех кроветворных клетках, но отсутствует
на зрелых Т-лимфоцитах; под влиянием интерлейкинов происходит экспрессия белка и
на этих клетках.
Роль la-белка в иммунном ответе сводится к следующему. АГ могут быть
распознаны иммунокомпетентными клетками лишь при контакте со специфическими
рецепторами, однако количество АГ слишком велико и природа не заготовила для них
соответствующего числа рецепторов, вот почему АГ («чужое») может быть узнан лишь
в комплексе со «своим», функцию которого и несет la-белок или антигены
HLA-DR.
Продукты деградации АГ, покинув макрофаг, частично вступают во взаимодействие
с la-белком, образуя с ним комплекс, стимулирующий деятельность
антигенпрезентирующей клетки. При этом макрофаг начинает секретировать ряд
интерлейкинов. ИЛ-1 действует на Т-амплифайер, в результате чего у
последнего появляется рецептор к комплексу la-белок + АГ. Именно эта реакция,
как и все последующие, обеспечивает специфичность иммунного ответа.
Активированный Т-амплифайер выделяет ИЛ-2, действующий на различные клоны
Т-хелперов и цитотоксические лимфоциты, принимающие участие в клеточном
иммунитете. Стимулированные клоны Т-хелперов секретируют ИЛ-3, ИЛ-4, ИЛ-5 и
ИЛ-6, оказывающие преимущественное влияние на эффекторное звено
иммунного ответа и тем самым способствующие переходу В-лимфоцитов в
антителопродуценты. Благодаря этому образуются AT, или иммуноглобулины.
Другие интерлейкины (ИЛ-7, ИЛ-9, ИЛ-10, ИЛ-12) влияют преимущественно на рост и
дифференцировку Т- и В-лимфоцитов и являются факторами надежности,
обеспечивающими иммунный ответ.
Клеточный иммунитет зависит от действия гуморальных факторов, выделяемых
цитотоксическими лимфоцитами (Т-киллерами). Эти соединения получили наименование
«перфорины» и «цитолизины».
Установлено, что каждый Т-эффектор способен лизировать несколько
чужеродных клеток-мишеней. Этот процесс осуществляется в три стадии: 1)
распознавание и контакт с клетками-мишенями; 2) летальный удар; 3) лизис
клетки-мишени. Последняя стадия не требует присутствия Т-эффектора, так как
осуществляется под влиянием перфоринов и цитолизинов. В стадию летального
удара перфорины и цитолизины действуют на мембрану клетки-мишени и образуют в
ней поры, через которые проникает вода, разрывающая клетки.
Среди гуморальных факторов, выделяемых в процессе иммунного ответа, следует
указать на фактор некроза опухолей (ФНО) и интерфероны.
Действие интерферонов неспецифично, так как они обладают различными
функциями — стимулируют деятельность НК-клеток и макрофагов, влияют
непосредственно на ДНК- и РНК-содержащие вирусы, подавляя их рост и активность,
задерживают рост и разрушают злокачественные клетки, возможно, за счет
усиления продукции ФНО (схема 6.1).
Гуморальный иммунный ответ обеспечивается AT, или иммуноглобулинами. У
человека различают пять основных классов иммуноглобулинов: IgA, IgG, IgM, IgE,
IgD. Все они имеют как общие, так и специфические детерминанты.
Иммуноглобулины класса G. У человека являются наиболее
важными. Концентрация IgG в крови достигает 9—18 г/л. Иммуноглобулины
класса G обеспечивают противоинфекционную защиту, связывают токсины, усиливают
фагоцитарную активность, активируют систему комплемента, вызывают
агглютинацию бактерий и вирусов, они способны переходить через плаценту,
обеспечивая новорожденному ребенку так называемый пассивный иммунитет. Это
означает, что если мать перенесла «детские инфекции» (корь, коклюш, скарлатина и
др.), то новорожденный ребенок в течение 3—6 мес. к этим заболеваниям
невосприимчив, так как содержит к возбудителям данных инфекций материнские
AT.
Иммуноглобулины класса А. Делят на две разновидности:
сывороточные и секреторные. Первые из них находятся в крови, вторые — в
различных секретах. Соответственно этому сывороточный IgA
принимает участие в общем иммунитете, а секреторный IgA
обеспечивает местный иммунитет, создавая барьер на пути
проникновения инфекций и токсинов в организм.
Секреторный IgA находится в наружных секретах — в слюне, слизи
трахеобронхиального дерева, мочеполовых путей, молоке, молозиве. Молекулы IgA,
присутствующие во внутренних секретах и жидкостях (синовиальная, амниотическая,
плевральная, цереброспинальная и др.), существенно отличаются от молекул
IgA, присутствующего в наружных секретах. Секреторный компонент, по всей
видимости, образуется в эпителиальных клетках и в дальнейшем присоединяется
к молекуле IgA.
IgA нейтрализуют токсины и вызывают агглютинацию микроорганизмов и
вирусов. Концентрация сывороточных IgA колеблется от 1,5 до 4,0 г/л.
Содержание IgA резко возрастает при заболеваниях верхних дыхательных путей,
пневмониях, инфекционных заболеваниях желудочно-кишечного тракта и др.
|
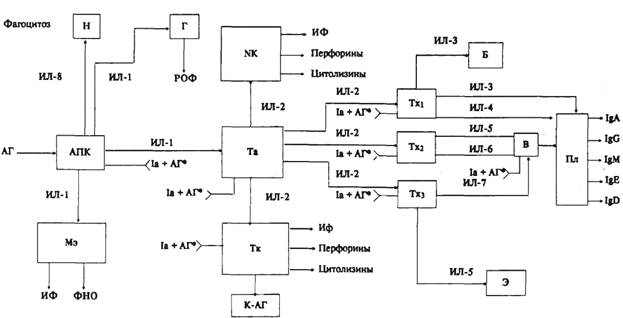
|
|
Схема 6.1. Иммунный ответ
AГ — антиген; AГ* — продукты деградации антигена; АПК —
антигенпрезентирующая клетка; Б — базофил; В — В-лимфоцит; Г — гепатоцит;
ИЛ — интерлейкин; ИФ — интерферон; К—АГ — чужеродная клетка; Мэ —
макрофаг-эффектор; Н — нейтрофил; Пл — плазматическая клетка; РОФ —
реактанты острой фазы воспаления; Та — Т-амплифайер; Тк — Т-киллер; Тх —
Т-хелпер; ФНО — фактор некроза опухолей; Э — эозинофил; 1а — la-подобный
белок; Ig — иммуноглобулин; NK — естественный киллер.
|
Иммуноглобулины класса IgE. Принимают участие в
нейтрализации токсинов, опсонизации, агглютинации и бактериолизисе,
осуществляемом комплементом. К этому классу также относятся некоторые
природные AT, например к чужеродным (не свойственным человеку) эритроцитам.
Содержание IgE повышается при инфекционных заболеваниях у взрослых и
детей.
Иммуноглобулины класса IgD. Обладают свойством
фиксироваться на базофилах и тучных клетках и вызывать в случае образования
иммунных комплексов их дегрануляцию. Содержание увеличивается при так называемых
аллергических заболеваниях — бронхиальной астме, вазомоторном рините,
гельминтозах, аллергических дерматитах и др.
Иммуноглобулины класса IgD. Представляют собой антитела,
локализующиеся в мембране плазматических клеток, в сыворотке концентрация их
невелика. Значение IgD не выяснено. Предполагают, что IgD принимает участие
в аутоиммунных процессах.
Регуляция иммунитета. Интенсивность иммунного ответа во многом
определяется состоянием нервной и эндокринной систем. Установлено, что
раздражение различных подкорковых структур (таламус, гипоталамус, серый бугор)
может сопровождаться как усилением, так и торможением иммунной реакции на
введение антигенов. Показано, что возбуждение симпатического отдела автономной
(вегетативной) нервной системы, как и введение адреналина, усиливает
фагоцитоз и интенсивность иммунного ответа. Повышение тонуса парасимпатического
отдела вегетативной нервной системы приводит к противоположным реакциям.
Стресс, а также депрессии угнетают иммунитет, что сопровождается не
только повышенной восприимчивостью к различным заболеваниям, но и создает
благоприятные условия для развития злокачественных новообразований.
За последние годы установлено, что гипофиз и эпифиз с помощью особых
пептидных биорегуляторов, получивших наименование «цитомедины», контролируют
деятельность тимуса. Передняя доля гипофиза является регулятором
преимущественно клеточного, а задняя — гуморального иммунитета.
Иммунная регуляторная система. В последнее время высказано
предположение, что существуют не две системы регуляции (нервная и гуморальная),
а три (нервная, гуморальная и иммунная). Иммунокомпетентные клетки способны
вмешиваться в морфогенез, а также регулировать течение физиологических функций.
Не подлежит сомнению, что Т-лимфоциты играют чрезвычайно важную роль в
регенерации тканей. Многочисленные исследования показывают, что Т-лимфоциты и
макрофаги осуществляют «хелперную» и «супрессорную» функции в отношении
эритропоэза и лейкопоэза. Лимфокины и монокины, выделяемые лимфоцитами,
моноцитами и макрофагами, способны изменять деятельность центральной нервной
системы, сердечно-сосудистой системы, органов дыхания и пищеварения,
регулировать сократительные функции гладкой и поперечнополосатой
мускулатуры.
Особенно важная роль в регуляции физиологических функций принадлежит
интерлейкинам, которые являются «семьей молекул на все случаи жизни», так как
вмешиваются во все физиологические процессы, протекающие в организме.
Иммунная система является регулятором гомеостаза. Эта функция осуществляется
за счет выработки аутоантител, связывающих активные ферменты, факторы
свертывания крови и избыток гормонов.
Иммунологическая регуляция, с одной стороны, является неотъемлемой
частью гуморальной, так как большинство физиологических и биохимических
процессов осуществляется при непосредственном участии гуморальных посредников.
Однако нередко иммунологическая регуляция носит прицельный характер и тем
самым напоминает нервную. Лимфоциты и моноциты, а также другие клетки,
принимающие участие в иммунном ответе, отдают гуморальный посредник
непосредственно органу-мишени. Отсюда предложение назвать иммунологическую
регуляцию клеточно-гуморальной. Основную роль в ней следует отвести различным
популяциям Т-лимфоцитов, осуществляющих «хелперные» и «супрессорные»
функции по отношению к различным физиологическим процессам.
Учет регуляторных функций иммунной системы позволяет врачам различных
специальностей по-новому подойти к решению многих проблем клинической
медицины.
6.2.3. Тромбоциты
Тромбоциты, или кровяные пластинки, образуются из гигантских клеток
красного костного мозга — мегакариоцитов. В костном мозге мегакариоциты
плотно прижаты к промежуткам между фибробластами и эндотелиальными клетками,
через которые их цитоплазма выдается наружу и служит материалом для образования
тромбоцитов. В кровотоке тромбоциты имеют круглую или слегка овальную форму,
диаметр их не превышает 2—3 мкм. У тромбоцита нет ядра, но имеется большое
количество гранул (до 200) различного строения. При соприкосновении с
поверхностью, отличающейся по своим свойствам от эндотелия, тромбоцит
активируется, распластывается и у него появляется до 10 зазубрин и
отростков, которые могут в 5—10 раз превышать диаметр тромбоцита. Наличие этих
отростков важно для остановки кровотечения.
В норме число тромбоцитов у здорового человека составляет 2—4-1011 /л, или
200—400 тыс. в 1 мкл. Увеличение числа тромбоцитов носит наименование
«тромбоцитоз», уменьшение — «тромбоцитопения». В естественных
условиях число тромбоцитов подвержено значительным колебаниям (количество их
возрастает при болевом раздражении, физической нагрузке, стрессе), но редко
выходит за пределы нормы. Как правило, тромбоцитопения является признаком
патологии и наблюдается при лучевой болезни, врожденных и приобретенных
заболеваниях системы крови.
Основное назначение тромбоцитов — участие в процессе гемостаза (см.
раздел 6.4). Важная роль в этой реакции принадлежит так называемым
тромбоцитарным факторам, которые сосредоточены главным образом в гранулах и
мембране тромбоцитов. Часть из них обозначают буквой Р (от слова platelet —
пластинка) и арабской цифрой (Р1, Р2 и т. д.). Наиболее
важными являются Р3, или частичный (неполный)
тромбопластин, представляющий осколок клеточной мембраны; Р4,
или антигепариновый фактор; Р5, или фибриноген тромбоцитов;
АДФ; контрактильный белок тромбастенин (напоминающий актомиозин),
вазоконстрикторные факторы — серотонин, адреналин, норадреналин и др.
Значительная роль в гемостазе отводится тромбоксану А2
(ТхА2), который синтезируется из арахидоновой кислоты, входящей
в состав клеточных мембран (в том числе и тромбоцитов) под влиянием фермента
тромбоксансинтетазы.
На поверхности тромбоцитов находятся гликопротеиновые образования,
выполняющие функции рецепторов. Часть из них «замаскирована» и
экспрессируется после активации тромбоцита стимулирующими агентами — АДФ,
адреналином, коллагеном, микрофибриллами и др.
Тромбоциты принимают участие в защите организма от чужеродных агентов.
Они обладают фагоцитарной активностью, содержат IgG, являются источником
лизоцима и β-лизинов, способных разрушать мембрану некоторых
бактерий. Кроме того, в их составе обнаружены пептидные факторы, вызывающие
превращение «нулевых» лимфоцитов (0-лимфоциты) в Т- и В-лимфоциты. Эти
соединения в процессе активации тромбоцитов выделяются в кровь и при травме
сосудов защищают организм от попадания болезнетворных микроорганизмов.
Регуляторами тромбоцитопоэза являются тромбоцитопоэтины кратковременного и
длительного действия. Они образуются в костном мозге, селезенке, печени, а также
входят в состав мегакариоцитов и тромбоцитов. Тромбоцитопоэтины
кратковременного действия усиливают отшнуровку кровяных пластинок от
мегакариоцитов и ускоряют их поступление в кровь; тромбоцитопоэтины
длительного действия способствуют переходу предшественников гигантских
клеток костного мозга в зрелые мегакариоциты. На активность тромбоцитопоэтинов
непосредственное влияние оказывают ИЛ-6 и ИЛ-11.
6.3. ГРУППЫ КРОВИ
6.3.1. Система АВО
Учение о группах крови возникло из потребностей клинической медицины.
Переливая кровь от животных человеку или от человека человеку, врачи нередко
наблюдали тяжелейшие осложнения, иногда заканчивавшиеся гибелью реципиента
(лицо, которому переливают кровь).
С открытием венским врачом К. Ландштейнером (1901) групп крови стало понятно,
почему в одних случаях трансфузии крови проходят успешно, а в других
заканчиваются трагически для больного. К. Ландштейнер впервые обнаружил,
что плазма, или сыворотка, одних людей способна агглютинировать (склеивать)
эритроциты других людей. Это явление получило наименование
изогемагглютинации. В основе ее лежит наличие в эритроцитах антигенов,
названных агглютиногенами и обозначаемых буквами А и В, а в плазме —
природных антител, или агглютининов, именуемых α и β.
Агглютинация эритроцитов наблюдается лишь в том случае, если встречаются
одноименные агглютиноген и агглютинин: А и α, В и β.
Установлено, что агглютинины, являясь природными антителами (AT), имеют два
центра связывания, а потому одна молекула агглютинина способна образовать
мостик между двумя эритроцитами. При этом каждый из эритроцитов может при
участии агглютининов связаться с соседним, благодаря чему возникает конгломерат
(агглютинат) эритроцитов.
В крови одного и того же человека не может быть одноименных агглютиногенов и
агглютининов, так как в противном случае происходило бы массовое склеивание
эритроцитов, что несовместимо с жизнью. Возможны только четыре комбинации, при
которых не встречаются одноименные агглютиногены и агглютинины, или четыре
группы крови: I — αβ, II — Aβ, III — Вα, IV —
АВ.
Кроме агглютининов, в плазме, или сыворотке, крови содержатся гемолизины:
их также два вида и они обозначаются, как и агглютинины, буквами α
и β. При встрече одноименных агглютиногена и гемолизина наступает
гемолиз эритроцитов. Действие гемолизинов проявляется при температуре 37—40
οС. Вот почему при переливании несовместимой крови у
человека уже через 30—40 с. наступает гемолиз эритроцитов. При комнатной
температуре, если встречаются одноименные агглютиногены и агглютинины,
происходит агглютинация, но не наблюдается гемолиз.
В плазме людей с II, III, IV группами крови имеются антиагглютиногены,
покинувшие эритроцит и ткани. Обозначаются они, как и агглютиногены, буквами А и
В (табл. 6.4).
Таблица 6.4. Серологический состав основных групп крови (система
АВО)
|
Группа крови |
Эритроциты |
Плазма, или сыворотка |
|
агглютиногены |
агглютинины и гемолизины |
антиагглютинины |
|
I(О) |
0 |
α,β |
0 |
|
II (А) |
А |
β |
А |
|
III (В) |
В |
а |
В |
|
IV (АВ) |
АВ |
|
АВ |
Как видно из приводимой таблицы, I группа крови не имеет агглютиногенов,
а потому по международной классификации обозначается как группа 0, II — носит
наименование А, III — В, IV — АВ.
Для решения вопроса о совместимости групп крови пользуются следующим
правилом: среда реципиента должна быть пригодна для жизни эритроцитов донора
(человек, который отдает кровь). Такой средой является плазма, следовательно, у
реципиента должны учитываться агглютинины и гемолизины, находящиеся в
плазме, а у донора — агглютиногены, содержащиеся в эритроцитах. Для решения
вопроса о совместимости групп крови смешивают исследуемую кровь с сывороткой,
полученной от людей с различными группами крови (табл. 6.5).
Таблица 6.5. Совместимость различных групп крови
|
Группа сыворотки
|
Группа эритроцитов |
|
I(О) |
II(A) |
III(В) |
IV (АВ) |
|
Iαβ |
- |
+ |
+ |
+ |
|
II β |
- |
- |
+ |
+ |
|
IIIα |
- |
+ |
- |
+ |
|
IV |
- |
- |
- |
- |
Примечание. «+» -- наличие агглютинации (группы несовместимы); «--» --
отсутствие агглютинации (группы совместимы.
 Из таблицы
видно, что агглютинация происходит в случае смешивания сыворотки I группы с
эритроцитами II, III и IV групп, сыворотки II группы — с эритроцитами III и IV
групп, сыворотки III группы с эритроцитами II и IV групп.
Из таблицы
видно, что агглютинация происходит в случае смешивания сыворотки I группы с
эритроцитами II, III и IV групп, сыворотки II группы — с эритроцитами III и IV
групп, сыворотки III группы с эритроцитами II и IV групп.
Следовательно, кровь I группы совместима со всеми другими группами крови,
поэтому человек, имеющий I группу крови, называется универсальным
донором. С другой стороны, эритроциты IV группы крови не должны давать
реакции агглютинации при смешивании с плазмой (сывороткой) людей с любой группой
крови, поэтому люди с IV группой крови называются универсальными
реципиентами.
Почему же при решении вопроса о совместимости не принимают в расчет
агглютинины и гемолизины донора? Это объясняется тем, что агглютинины и
гемолизины при переливании небольших доз крови (200—300 мл) разводятся в большом
объеме плазмы (2500— 2800 мл) реципиента и связываются его антиагглютининами, а
потому не должны представлять опасности для эритроцитов.
В повседневной практике для решения вопроса о группе переливаемой крови
пользуются иным правилом: переливаться должны одногруппная кровь и только по
жизненным показаниям, когда человек потерял много крови. Лишь в случае
отсутствия одногруппной крови с большой осторожностью можно перелить небольшое
количество иногруппной совместимой крови. Объясняется это тем, что
приблизительно у 10—20% людей имеется высокая концентрация очень активных
агглютининов и гемолизинов, которые не могут быть связаны антиагглютининами даже
в случае переливания небольшого количества иногруппной крови.
Посттрансфузионные осложнения иногда возникают из-за ошибок при определении
групп крови. Установлено, что агглютиногены А и В существуют в разных вариантах,
различающихся по своему строению и антигенной активности. Большинство из них
получило цифровое обозначение (А1, А,2, А3 и т.
д., В1, В2 и т. д.). Чем больше порядковый номер
агглютиногена, тем меньшую активность он проявляет. И хотя разновидности
агглютиногенов А и В встречаются относительно редко, при определении групп крови
они могут быть не обнаружены, что может привести к переливанию несовместимой
крови.
Следует также учитывать, что большинство человеческих эритроцитов несет
антиген Н. Этот АГ всегда находится на поверхности клеточных мембран у лиц с
группой крови 0, а также присутствует в качестве скрытой детерминанты на клетках
людей с группами крови А, В и АВ. Н — антиген, из которого образуются антигены А
и В. У лиц с I группой крови антиген доступен действию анти-Н-антител, которые
довольно часто встречаются у людей со II и IV группами крови и относительно
редко у лиц с III группой. Это обстоятельство может послужить причиной
гемотрансфузионных осложнений при переливании крови 1 группы людям с
другими группами крови.
Концентрация агглютиногенов на поверхности мембраны эритроцитов
чрезвычайно велика. Так, один эритроцит группы крови A1 содержит в
среднем 900 000—1 700 000 антигенных детерминант, или рецепторов, к одноименным
агглютининам. С увеличением порядкового номера агглютиногена число таких
детерминант уменьшается. Эритроцит группы А2 имеет всего 250
000—260 000 антигенных детерминант, что также объясняет меньшую активность
этого агглютиногена.
В настоящее время система AB0 часто обозначается как АВН, а вместо терминов
«агглютиногены» и «агглютинины» применяются термины «антигены» и «антитела»
(например, АВН-антигены и АВН-антитела).
6.3.2. Система резус (Rh-hr) и другие
К. Ландштейнер и А. Винер (1940) обнаружили в эритроцитах обезьяны макаки
резус АГ, названный ими резус-фактором. В дальнейшем оказалось, что
приблизительно у 85% людей белой расы также имеется этот АГ. Таких людей
называют резус-положительными (Rh+). Около 15% людей этот АГ не
имеют и носят название резус-отрицательных (Rh).
Известно, что резус-фактор — это сложная система, включающая более 40
антигенов, обозначаемых цифрами, буквами и символами. Чаще всего встречаются
резус-антигены типа D (85%), С (70%), Е (30%), е (80%) — они же и обладают
наиболее выраженной антигенностью. Система резус не имеет в норме одноименных
агглютининов, но они могут появиться, если резус-отрицательному человеку
перелить резус-положительную кровь.
Резус-фактор передается по наследству. Если женщина Rh, a мужчина
Rh+, то плод в 50—100% случаев унаследует резус-фактор от отца, и
тогда мать и плод будут несовместимы по резус-фактору. Установлено, что при
такой беременности плацента обладает повышенной проницаемостью по отношению
к эритроцитам плода. Последние, проникая в кровь матери, приводят к образованию
антител (антирезусагглютининов). Проникая в кровь плода, антитела вызывают
агглютинацию и гемолиз его эритроцитов.
Тяжелейшие осложнения, возникающие при переливании несовместимой крови и
резус-конфликте, обусловлены не только образованием конгломератов
эритроцитов и их гемолизом, но и интенсивным внутрисосудистым свертыванием
крови, так как в эритроцитах содержится набор факторов, вызывающих
агрегацию тромбоцитов и образование фибриновых сгустков. При этом страдают все
органы, но особенно сильно повреждаются почки, так как сгустки забивают
«чудесную сеть» клубочка почки, препятствуя образованию мочи, что может быть
несовместимо с жизнью.
Согласно современным представлениям, мембрана эритроцита рассматривается как
набор самых различных АГ, которых насчитывается более 500. Только из этих
АГ можно составить более 400 млн. комбинаций, или групповых признаков крови.
Если же учитывать и все остальные АГ, встречающиеся в крови, то число комбинаций
достигнет 700 млрд., т. е. значительно больше, чем людей на земном шаре.
Разумеется, далеко не все АГ важны для клинической практики. Однако при
переливании крови со сравнительно редко встречающимся АГ могут возникнуть
тяжелейшие гемотрансфузионные осложнения и даже смерть больного.
Нередко при беременности возникают серьезные осложнения, в том числе
выраженная анемия, что может быть объяснено несовместимостью групп крови по
системам мало изученных антигенов матери и плода. При этом страдает не только
беременная, но в неблагополучных условиях находится и будущий ребенок.
Несовместимость матери и плода по группам крови может быть причиной
выкидышей и преждевременных родов.
Гематологи выделяют наиболее важные антигенные системы: ABO, Rh, MNSs, P,
Лютеран (Lu), Келл-Келлано (Kk), Льюис (Le), Даффи (Fy) и Кид (Jk). Эти системы
антигенов учитываются в судебной медицине для установления отцовства и иногда
при трансплантации органов и тканей.
В настоящее время переливание цельной крови производится сравнительно редко,
так как пользуются трансфузией различных компонентов крови, т. е. переливают то,
что больше всего требуется организму: плазму или сыворотку, эритроцитную,
лейкоцитную или тромбоцитную массу. В подобной ситуации вводится меньшее
количество антигенов, что снижает риск посттрансфузионных
осложнений.
6.3.3. Группы крови и
заболеваемость
Люди, имеющие различные группы крови, в неодинаковой мере подвержены тем или
иным заболеваниям. Так, у людей с I (0) группой крови чаше встречается язвенная
болезнь желудка и двенадцатиперстной кишки. Эти факты объясняются тем, что
агглютиногены А и В, выделяясь в составе желудочного и поджелудочного сока,
предохраняют стенку от повреждения протеолитическими ферментами. Люди, имеющие
II (А) группу крови, чаще страдают и тяжелее переносят сахарный диабет, у них
повышена свертываемость крови, из-за чего возникают инфаркты миокарда и
инсульты. Согласно статистическим данным, у лиц с II (А) группой крови чаще
встречаются раковые заболевания желудка и половых органов, а у лиц III (В)
группы — рак толстой кишки. Вместе с тем лица, имеющие I и IV группы крови,
менее восприимчивы к возбудителям чумы, но у них тяжелее протекает натуральная
оспа.
У резус-отрицательных людей различные заболевания крови встречаются
приблизительно в 6 раз чаще, чем у резус-положительных.
6.3. СИСТЕМА ГЕМОСТАЗА
Под термином «гемостаз» понимают комплекс реакций, направленных на остановку
кровотечения при травме сосудов. Значение системы гемостаза намного сложнее и
шире. Факторы гемостаза принимают участие в сохранении жидкого состояния крови,
регуляции транскапиллярного обмена, резистентности сосудистой стенки,
влияют на интенсивность репаративных процессов и др.
Принято различать сосудисто-тромбоцитарный гемостаз и процесс свертывания
крови. В первом случае речь идет об остановке кровотечения из мелких
сосудов с низким кровяным давлением, диаметр которых не превышает 100 мкм, во
втором — о борьбе с кровопотерей при повреждениях артерий и вен. Такое деление
носит условный характер, потому что при повреждении как мелких, так и крупных
кровеносных сосудов всегда наряду с образованием тромбоцитарной пробки
осуществляется свертывание крови.
6.4.1. Сосудисто-тромбоцитарный
гемостаз
Сосудисто-тромбоцитарный гемостаз сводится к образованию тромбоцитарной
пробки, или тромбоцитарного тромба. Условно его разделяют на три стадии: 1)
временный (первичный) спазм сосудов; 2) образование тромбоцитарной пробки за
счет адгезии (прикрепления к поврежденной поверхности) и агрегации
(склеивания между собой) тромбоцитов; 3) ретракция (сокращение и уплотнение)
тромбоцитарной пробки.
Сразу после травмы наблюдается первичный спазм кровеносных сосудов,
благодаря чему кровотечение в первые секунды может не возникнуть или носит
ограниченный характер. Первичный спазм сосудов обусловлен выбросом в кровь в
ответ на болевое раздражение адреналина и норадреналина и длится не более 10—15
с. В дальнейшем наступает вторичный спазм, обусловленный активацией
тромбоцитов и отдачей в кровь сосудосуживающих агентов — серотонина,
ТхА2, адреналина и др.
Повреждение сосудов сопровождается немедленной активацией тромбоцитов, что
обусловлено появлением высоких концентраций АДФ (из разрушающихся эритроцитов и
травмированных сосудов), а также с обнажением субэндотелия, коллагеновых и
фибриллярных структур. В результате «раскрываются» вторичные рецепторы и
создаются оптимальные условия для адгезии, агрегации и образования
тромбоцитарной пробки.
Адгезия обусловлена наличием в плазме и тромбоцитах особого белка — фактора
Виллебранда (FW), имеющего три активных центра, два из которых связываются
с экспрессированными рецепторами тромбоцитов, а один — с рецепторами
субэндотелия и коллагеновых волокон. Таким образом, тромбоцит с помощью FW
оказывается «подвешенным» к травмированной поверхности сосуда.
Одновременно с адгезией наступает агрегация тромбоцитов, осуществляемая
с помощью фибриногена — белка, содержащегося в плазме и тромбоцитах и
образующего между ними связующие мостики, что и приводит к появлению
тромбоцитарной пробки.
Важную роль в адгезии и агрегации играет комплекс белков и полипептидов,
получивших наименование «интегрины». Последние служат связующими агентами между
отдельными тромбоцитами (при склеивании друг с другом) и структурами
поврежденного сосуда. Агрегация тромбоцитов может носить обратимый характер
(вслед за агрегацией наступает дезагрегация, т. е. распад агрегатов), что
зависит от недостаточной дозы агрегирующего (активирующего) агента.
Из тромбоцитов, подвергшихся адгезии и агрегации, усиленно секретируются
гранулы и содержащиеся в них биологически активные соединения — АДФ,
адреналин, норадреналин, фактор Р4, ТхА2 и др. (этот
процесс получил название реакции высвобождения), что приводит к вторичной,
необратимой агрегации. Одновременно с высвобождением тромбоцитарных факторов
происходит образованием тромбина, резко усиливающего агрегацию и
приводящего к появлению сети фибрина, в которой застревают отдельные
эритроциты и лейкоциты.
Благодаря контрактильному белку тромбостенину тромбоциты подтягиваются друг к
другу, тромбоцитарная пробка сокращается и уплотняется, т. е. наступает ее
ретракция.
В норме остановка кровотечения из мелких сосудов занимает 2—4 мин.
Важную роль для сосудисто-тромбоцитарного гемостаза играют производные
арахидоновой кислоты — простагландин I2 (PgI2), или
простациклин, и ТхА2. При сохранении целости эндотелиального покрова
действие Pgl преобладает над ТхА2, благодаря чему в сосудистом русле
не наблюдается адгезии и агрегации тромбоцитов. При повреждении эндотелия в
месте травмы синтез Pgl не происходит, и тогда проявляется влияние
ТхА2, приводящее к образованию тромбоцитарной пробки.
6.4.2. Процесс свертывания
крови
При повреждении крупных кровеносных сосудов (артерий, вен, артериол), также
происходит образование тромбоцитарной пробки, но она неспособна остановить
кровотечение, так как легко вымывается током крови. Основное значение в
этом процессе принадлежит свертыванию крови, сопровождающемуся в конечном итоге
образованием плотного фибринового сгустка.
6.4.2.1. Плазменные и
клеточные факторы свертывания крови
В свертывании крови принимает участие комплекс белков, находящихся в
плазме (плазменные факторы гемокоагуляции), большинство из которых является
проферментами. В отличие от тромбоцитарных факторов они обозначаются римскими
цифрами.
ПЛАЗМЕННЫЕ ФАКТОРЫ СВЕРТЫВАНИЯ КРОВИ
|
I, или фибриноген
|
Белок. Образуется в печени. Под влиянием тромбина переходит в фибрин.
Участвует в агрегации тромбоцитов. Необходим для репарации тканей
|
|
II, или протромбин |
Гликопротеид. Образуется в печени в присутствии витамина К. Под
влиянием протромбиназы переходит в тромбин (фактор Ив)
|
|
Ш, или тромбопластин
|
Состоит из белка апопротеина III и комплекса фосфолипидов. Входит в
состав мембран многих тканей. Является матрицей для развертывания
реакций, направленных на образование протромбиназы по внешнему
механизму |
|
IV, или ион Са2*
|
Участвует в образовании комплексов, входит в состав протромбиназы.
Способствует агрегации тромбоцитов. Связывает гепарин. Принимает
участие в ретракции сгустка и тромбоцитарной пробки. Тормозит
фибринолиз |
|
V, или акцелератор-глобулин
|
Белок. Образуется в печени. Активизируется тромбином (фактор Па).
Создает оптимальные условия для взаимодействия фактора Ха и протромбина
(фактор II) |
|
(VI, исключен из классификации)
|
|
|
VII, или проконвертин
|
Гликопротеид. Образуется в печени под влиянием витамина К. Принимает
участие в формировании протромбиназы по внешнему механизму. Активируется
факторами Х11в, Ха, 1Ха, Па и при взаимодействии с тромбопластином (фактор
III) |
|
VIII, или антигемофильный глобулин (АГГ). антигемофильный глобулин
А
|
Гликопротеид. Синтезируется в печени, селезенке, лейкоцитах. Образует
комплексную молекулу с фактором Виллебранда (FW) и специфическим
антигеном. Активируется тромбином. Создает оптимальные условия для
взаимодействия факторов 1Ха и X. При его отсутствии возникает заболевание
гемофилия А
|
|
VIII: FW
|
Компонент комплекса фактора VIII: Образуется эндотелиальными клетками.
Обеспечивает устойчивость фактора VIII: С в кровотоке и необходим для
адгезии тромбоцитов. При его недостатке развивается болезнь Виллебранда,
сопровождающаяся нарушением сосудисто-тромбоцитарного гемостаза |
|
IX, или Кристмас-фактор,
антигемофильный фактор
В
|
Гликопротеид. Образуется в печени под влиянием витамина К. Активируется
факторами XIa, VIla и IIа. Переводит фактор X в Ха. При его отсутствии
возникает заболевание гемофилия В |
|
X, или Стюарт Прауэр-фактор
|
Гликопротеид. Образуется в печени, под влиянием витамина К. Фактор Ха,
являясь протромбиназой, активируется факторами VIla и IХа. Переводит
фактор II в IIа
|
|
XI, или плазменный предшественник тромбопластина
|
Гликопротеид. Предполагают, что образуется в печени. Активируется
фактором ХIIа калликреином совместно с высокомолекулярным кининогеном
(ВМК) |
|
XII, или фактор Хагемана
|
Белок. Предполагают, что образуется эндотелиальными клетками,
лейкоцитами, макрофагами. Активируется отрицательно заряженными
поверхностями, адреналином, калликреином. Запускают внешний и
внутренний механизм образования протромбиназы и фибринолиза,
активирует фактор XI и прекалликреин |
|
ХШ, или фибринстабилизирующий фактор (ФСФ), фибриназа
|
Глобулин. Синтезируется фибробластами и мегакариоцитами. Стабилизирует
фибрин. Необходим для нормального течения репаративных процессов |
|
Фактор Флетчера, или прекалликреин
Является компонентом калликреин-кининовой системы.
|
Белок. Участвует в активации фактора XII, плазминогена и ВМК |
|
Фактор Фитцджеральда, высокомолекулярный кининоген (ВМК)
Является компонентом калликреин-кининовой системы.
|
Образуется в тканях. Активируется калликреином, принимает участие в
активации факторов XII, XI и фибринолиза |
Активация плазменных факторов происходит главным образом за счет протеолиза и
сопровождается отщеплением пептидных ингибиторов. Активное состояние
фактора обозначается присоединением к его номеру буквы «а» (фактор IIа, Va,
VIIa и т. д.). Плазменные факторы делят на 2 группы: витамин К-зависимые
(образуются преимущественно в печени под влиянием витамина К) и витамин
К-независимые (для синтеза которых витамин К не требуется).
В эритроцитах обнаружены многие соединения, аналогичные тромбоцитарным
факторам (см. раздел 6.2.3). Важнейшим из них является фосфолипидный фактор, или
частичный тромбопластин (напоминает фактор Р3), который входит в
состав мембраны. Кроме того, эритроциты содержат большое количество АДФ,
фибриназу и другие факторы. При травме сосуда около 1% наименее стойких
эритроцитов вытекающей крови разрушается, что способствует образованию
тромбоцитарной пробки и фибринового сгустка.
Особенно велика роль эритроцитов в свертывании крови в случае их массового
разрушения (переливание несовместимой крови, резус-конфликт матери и плода,
гемолитические анемии и др.)
Лейкоциты содержат факторы свертывания, получившие наименование
лейкоцитарных. В частности, моноциты и макрофаги при стимуляции антигеном
синтезируют белковую часть тромбопластина — апопротеин III, что значительно
ускоряет свертывание крови. Эти же клетки являются продуцентами витамин
К-зависимых факторов свертывания — II, VII, IX и X. Приведенные факторы
являются одной из основных причин возникновения диссеминированного
(распространенного) внутрисосудистого свертывания крови (ДВС-синдром) при
многих воспалительных и инфекционных заболеваниях, что значительно отягощает
течение патологического процесса, а иногда служит причиной смерти больных.
Важная роль в процессе свертывания крови отводится тканевым факторам, к
которым в первую очередь относится тромбопластин (фактор 3). Концентрация
тромбопластина высока в коре большого мозга, легких, плаценте и стимулированном
антигенами эндотелии сосудов. При разрушении тканей и стимуляции эндотелия
большое количество тромбопластина поступает в кровоток, что может вызывать
развитие ДВС-синдрома.
6.4.2.2. Механизм свертывания
крови
Процесс свертывания крови представляет собой преимущественно
проферментно-ферментный каскад, в котором проферменты, переходя в активное
состояние, приобретают способность активировать другие факторы свертывания
крови. Подобная активация может носить последовательный и ретроградный
характер.
Процесс свертывания крови может быть разделен на три фазы: первая включает
комплекс последовательных реакций, приводящих к образованию протромбиназы, во
вторую фазу осуществляется переход протромбина (фактор II) в тромбин (фактор
IIа) и в третью фазу из фибриногена образуется фибрин.
Первая фаза — образование протромбиназы может происходить по внешнему и
внутреннему механизму. Внешний механизм предполагает обязательное
присутствие тромбопластина (фактор III), внутренний же связан с участием
тромбоцитов (фактор Р3) или разрушенных эритроцитов. Вместе с тем внутренний и
внешний пути образования протромбиназы имеют много общего, так как активируются
одними и теми же факторами (фактор ХIIа, калликреин, ВМК и др.), а также
приводят в конечном итоге к появлению одного и того же активного фермента —
фактора Ха, выполняющего функции протромбиназы. При этом и полный, и
частичный тромбопластин служат матрицами, на которых в присутствии ионов
Са2+ развертываются ферментативные реакции.
Формирование протромбиназы по внешнему пути начинается с активации фактора
VII при его взаимодействии с тромбопластином и фактором ХIIа. Кроме того, фактор
VII может переходить в деятельное состояние под влиянием факторов XIa, IXa, Ха,
IIа и калликреина. В свою очередь фактор VIIa не только переводит фактор X в Ха
(ведет к появлению протромбиназы), но и активирует фактор IX, участвующий в
образовании протромбиназы по внутреннему механизму.
Образование протромбиназы по внешнему пути происходит чрезвычайно быстро (за
20—30 с), ведет к появлению небольших порций тромбина (IIа), который
способствует необратимой агрегации тромбоцитов, активации факторов VIII и V
и значительно ускоряет формирование протромбиназы по внутреннему механизму.
Инициатором внутреннего механизма образования протромбиназы является фактор XII,
который активируется травмированной поверхностью стенки сосуда, кожей,
коллагеном, адреналином, в лабораторных условиях — при контакте со стеклом,
после чего переводит фактор XI в XIa. В этой реакции может принимать участие
калликреин (активируется фактором ХIIа) и ВМК (активируется калликреином).
Фактор XIa оказывает непосредственное влияние на фактор IX, переводя его в
фактор IXa. Специфическая деятельность последнего направлена на протеолиз
фактора X и протекает при обязательном участии фактора VIII (или
VIIIa).
Следует заметить, что активация фактора X под влиянием комплекса факторов
VIII и IXa получила название теназной реакции.
Вторая фаза процесса свертывания крови — переход фактора II в фактор IIа
осуществляется под влиянием протромбиназы (фактор Ха) в присутствии фактора
V (Va) и сводится к протеолитическому расщеплению протромбина, благодаря чему
появляется фермент тромбин, обладающий свертывающей активностью.
Третья стадия процесса свертывания крови — переход фибриногена в фибрин —
носит этапный характер. Под влиянием фактора IIа от фибриногена отщепляются
фибринопептиды и образуется фибрин-мономер (фактор Im). Из него благодаря
процессу полимеризации формируются олигомеры и димеры фибрина (фактор Iо и Id),
из которых за счет продольного и поперечного связывания образуются протофибриллы
— легкорастворимый фибрин, или фибрин S, быстро лизирующийся под влиянием
протеаз (плазмина, трипсина). В дальнейшем в процесс образования фибрина
вмешивается фактор XIII (фибриназа, фибринстабилизирующий фактор), который после
активации тромбином в присутствии ионов Са2+
«прошивает» фибринполимеры дополнительными перекрестными связями, в результате
чего появляется труднорастворимый фибрин, или фибрин i (insoluble). В
результате этой реакции сгусток становится резистентным к фибринолитическим
(протеолитическим) агентам и плохо поддается разрушению (схема 6.2).
|
|
|
Схема 6.2. Процесс свертывания крови
Примечание. Для упрощения на схеме не указана ретроградная
активация факторов V, VII, VIII. Сплошная линия — переход фактора в
активное состояние, прерывистая — активация факторов свертывания.
|
Образовавшийся фибриновый сгусток благодаря тромбоцитам, входящим в его
структуру, сокращается и уплотняется (наступает ретракция) и прочно закупоривает
поврежденный сосуд.
6.4.3. Естественные
антикоагулянты
Несмотря на то что в циркулирующей крови имеются все факторы,
необходимые для образования тромба, в естественных условиях при наличии
целостности сосудов кровь остается жидкой. Это обусловлено наличием в
кровотоке противосвертывающих веществ, получивших название естественных
антикоагулянтов, или фибринолитического звена системы гемостаза.
Естественные антикоагулянты делят на первичные и вторичные. Первичные
антикоагулянты всегда присутствуют в циркулирующей крови, вторичные — образуются
в результате протеолитического расщепления факторов свертывания крови в процессе
образования и растворения фибринового сгустка.
Первичные антикоагулянты можно разделить на три основные группы: 1)
антитромбопластины — обладающие антитромбопластическим и антипротромбиназным
действием; 2) антитромбины — связывающие тромбин; 3) ингибиторы самосборки
фибрина — дающие переход фибриногена в фибрин.
Следует заметить, что при снижении концентрации первичных естественных
антикоагулянтов создаются благоприятные условия для развития тромбозов и
ДВС-синдрома.
ОСНОВНЫЕ ЕСТЕСТВЕННЫЕ АНТИКОАГУЛЯНТЫ (по Баркагану 3. С. и
Бишевскому К. М.)
|
Первичные |
|
Антитромбин III |
γ2-Глобулин. Синтезируется в печени. Прогрессивно
действующий ингибитор тромбине, факторов Ха, IXa, XIa, ХIIa, калликреина и
в меньшей степени — плазмина и трипсина. Плазменный кофактор
гепарина |
|
Гепарин |
Сульфатированный полисахарид. Трансформирует
антитромбин III из прогрессивного в антикоагулянт немедленного
действия, значительно повышая его активность. Образует с
тромбогенными белками и гормонами комплексы, обладающие
антикоагулянтным и неферментным фибринолитическим действием |
|
α2-Антиплазмнн |
Белок. Ингибирует действие плазмина, трипсина,
химотрипсина, калликреина, фактора Ха, урокиназы |
|
α2-Макроглобулин |
Прогрессивный ингибитор тромбина, калликреина,
плазмина и трипсина |
|
α2-Антитрипсин |
Ингибитор тромбина, трипсина и плазмина |
|
C1-эстеразный ингибитор |
α2-Нейроаминогликопротеид. Инактивирует калликреин,
предотвращая его действие на кининоген, факторы ХIIа, IXa, XIa и
плазмин |
|
Липопротеин-ассоциированный коагуляционный ингибитор (ЛАКИ) |
Ингибирует комплекс тромбопластин—фактор VII, инактивирует фактор
Ха |
|
Аполипопротеин А-11 |
Ингибирует комплекс тромбопластин—фактор VII |
|
Плацентарный антикоагулянтный протеин |
Образуется в плаценте. Ингибирует комплекс тромбопластин—фактор
VII |
|
Протеин С |
Витамин К-зависимый белок. Образуется в печени и в эндотелии. Обладает
свойствами сериновой протеазы. Вместе с протеином S связывает факторы Va и
VIIIa и активирует фибринолиз |
|
Протеин S |
Витамин К-зависимый белок, образуется эндотелиальными клетками.
Усиливает действие протеина С |
|
Тромбомодулин |
Кофактор протеина С, связывается с фактором IIa Образуется
эндотелиальными клетками |
|
Ингибитор самосборки фибрина |
Полипептид, образуется в различных тканях. Действует на фибрин-мономер
и полимер |
|
«Плавающие» рецепторы |
Гликопротеиды, связывают факторы IIа и Ха, а возможно, и другие
сериновые протеазы |
|
Аутоантитела к активным факторам свертывание |
Находятся в плазме, ингибируют факторы IIа, Ха и др. |
|
Вторичные
(образуются в процессе протеолиза — при свертывании крови, фибринолизе
и т. д.) |
|
Антитромбин I |
Фибрин. Адсорбирует и инактивирует тромбин |
|
Дериваты (продукты деградации) протромбина Р, R, Q и др. |
Ингибируют факторы Ха, Va |
|
Метафактор Va |
Ингибитор фактора Ха |
|
Метафактор ХIa |
Ингибитор комплекса ХIIа+Х1а |
|
Фибринопептиды |
Продукты протеолиза фибриногена тромбином; ингибируют фактор
IIа |
|
Продукты деградации фибриногена и фибрина (чаще последнего)
(ПДФ) |
Нарушают полимеризацию фибрин-мономера, блокируют фибриноген и
фибрин-мономер (образуют с ними комплексы), ингибируют факторы ХIа, IIа,
фибринолиз и агрегацию тромбоцитов |
К вторичным антикоагулянтам относят «отработанные» факторы
свертывания крови (принявшие участие в свертывании) и продукты деградации
фибриногена и фибрина (ПДФ), обладающие мощным антиагрегационным и
противосвертывающим действием, а также стимулирующие фибринолиз. Роль вторичных
антикоагулянтов сводится к ограничению внутрисосудистого свертывания крови
и распространения тромба по сосудам.
6.4.4. Фибринолиз
Фибринолиз является неотъемлемой частью системы гемостаза, всегда
сопровождает процесс свертывания крови и активируется факторами,
принимающими участие в этом процессе. Являясь важной защитной реакцией,
фибринолиз предотвращает закупорку кровеносных сосудов фибриновыми сгустками.
Кроме того, фибринолиз ведет к реканализации сосудов после остановки
кровотечения.
Ферментом, разрушающим фибрин, является плазмин (иногда его называют
«фибринолизин»), который в циркуляции находится в неактивном состоянии в виде
профермента плазминогена.
Фибринолиз, как и процесс свертывания крови, может протекать по внешнему и
внутреннему механизму (пути). Внешний механизм активации фибринолиза
осуществляется при участии тканевых активаторов, которые синтезируются
главным образом в эндотелии сосудов. К ним относятся тканевый активатор
плазминогена (ТАП) и урокиназа. Последняя также образуется в юкстагломеруляриом
комплексе (аппарате) почки (см. главу 12). Внутренний механизм активации
фибринолиза осуществляется плазменными активаторами, а также
активаторами форменных элементов крови — лейкоцитов, тромбоцитов и
эритроцитов и разделяется на Хагеман-зависимый и Хагеман-независимый.
Хагеман-зависимый фибринолиз протекает под влиянием факторов XIIа, калликреина и
ВМК, которые переводят плазминоген в плазмин. Хагеман-независимый
фибринолиз осуществляется наиболее быстро и носит срочный характер. Его
основное назначение сводится к очищению сосудистого русла от
нестабилизированного фибрина, образующегося в процессе внутрисосудистого
свертывания крови.
Образовавшийся в результате активации плазмин вызывает расщепление
фибрина (схема 6.3). При этом появляются ранние (крупномолекулярные) и
поздние (низкомолекулярные) ПДФ.
|
|
|
Схема 6.3. Фибринолиз
|
В плазме находятся и ингибиторы фибринолиза. Важнейшими из них являются
α2-антиплазмин, связывающий плазмин, трипсин, калликреин, урокиназу,
ТАП и, следовательно, вмешивающийся в процесс фибринолиза как на ранних, так и
на поздних стадиях. Сильным ингибитором плазмина служит α1-протеазный
ингибитор. Кроме того, фибринолиз тормозится α2-макроглобулином,
C1-протеазным ингибитором, а также рядом ингибиторов активатора
плазминогена, синтезируемых эндотелием, макрофагами, моноцитами и
фибробластами.
Фибринолитическая активность крови во многом определяется соотношением
активаторов и ингибиторов фибринолиза.
При ускорении свертывания крови и одновременном торможении фибринолиза
создаются благоприятные условия для развития тромбозов, эмболии и
ДВС-синдрома.
Наряду с ферментативным фибринолизом, по мнению профессора Б. А.
Кудряшова, существует так называемый неферментативный фибринолиз, который
обусловлен комплексными соединениями естественного антикоагулянта гепарина
с ферментами и гормонами. Неферментативный фибринолиз приводит к расщеплению
нестабилизированного фибрина, очищая сосудистое русло от фибрин-мономеров и
фибрина s.
6.4.5. Регуляция свертывания крови и фибринолиза
Свертывание крови, контактирующей с травмированными тканями,
осуществляется за 5—10 мин. Основное время в этом процессе уходит на образование
протромбиназы, тогда как переход протромбина в тромбин и фибриногена в
фибрин осуществляется довольно быстро. В естественных условиях время свертывания
крови может уменьшаться (развивается гиперкоагуляция) или удлиняться
(возникает гипокоагуляция).
Значительный вклад в изучение регуляции свертывания крови и фибринолиза
внесли отечественные ученые Е. С. Иваницкий-Василенко, А. А. Маркосян, Б. А.
Кудряшов, С. А. Георгиева и др.
Установлено, что при острой кровопотере, гипоксии, интенсивной мышечной
работе, болевом раздражении, стрессе свертывание крови значительно ускоряется,
что может привести к появлению фибрин-мономеров и даже фибрина s в сосудистом
русле. Однако благодаря одновременной активации фибринолиза, носящего защитный
характер, появляющиеся сгустки фибрина быстро растворяются и не наносят
вреда здоровому организму.
Ускорение свертывания крови и усиление фибринолиза при всех перечисленных
состояниях обусловлены повышением тонуса симпатической части автономной
нервной системы и поступлением в кровоток адреналина и норадреналина. При этом
активируется фактор Хагемана, что приводит к запуску внешнего и внутреннего
механизма образования протромбиназы, а также стимуляции Хагеман-зависимого
фибринолиза. Кроме того, под влиянием адреналина усиливается образование
апопротеина III — составной части тромбопластина, и наблюдается отрыв клеточных
мембран от эндотелия, обладающих свойствами тромбопластина, что способствует
резкому ускорению свертывания крови. Из эндотелия также выделяются ТАП и
урокиназа, приводящие к стимуляции фибринолиза (схема 6.4).
|
|
|
Схема 6.4. Влияние автономной нервной системы на свертывание
крови и фибринолиз
Примечание. Для упрощения опущены промежуточные этапы
образования тромбина.
|
В случае повышения тонуса парасимпатической части автономной нервной системы
(раздражение блуждающего нерва, введение АХ, пилокарпина) также наблюдаются
ускорение свертывания крови и стимуляция фибринолиза. В этих условиях происходит
выброс тромбопластина и активаторов плазминогена из эндотелия сердца и
сосудов. Следовательно, основным эфферентным регулятором свертывания крови и
фибринолиза является сосудистая стенка. Напомним также, что в эндотелии
сосудов синтезируется Pgl2, препятствующий в кровотоке адгезии и
агрегации тромбоцитов. Вместе с тем развивающаяся гиперкоагуляция может
смениться гипокоагуляцией, которая в естественных условиях носит вторичный
характер и обусловлена расходом (потреблением) тромбоцитов и плазменных факторов
свертывания крови, образованием вторичных антикоагулянтов, а также
рефлекторным выбросом в сосудистое русло в ответ на появление фактора IIа,
гепарина и антитромбина III (см. схему 6.4).
При многих заболеваниях, сопровождающихся разрушением эритроцитов,
лейкоцитов, тромбоцитов и тканей и/или гиперпродукцией апопротеина III
стимулированными эндотелиальными клетками, моноцитами и макрофагами (эта
реакция опосредована действием антигенов и интерлейкинов), развивается
ДВС-синдром, значительно отягощающий течение патологического
процесса и даже приводящий к смерти больного. В настоящее время ДВС-синдром
обнаружен более чем при 100 различных заболеваниях. Особенно часто он возникает
при переливании несовместимой крови, обширных травмах, отморожениях,
ожогах, длительных оперативных вмешательствах на легких, печени, сердце,
предстательной железе, всех видах шока, краш-синдроме (длительное сдавление
конечностей), а также в акушерской практике при попадании в кровоток матери
околоплодных вод, насыщенных тромбопластином плацентарного происхождения.
При этом возникает гиперкоагуляция, которая из-за интенсивного потребления
тромбоцитов, фибриногена, факторов V, VIII, XIII и др. в результате интенсивного
внутрисосудистого свертывания крови сменяется вторичной гипокоагуляцией вплоть
до полной неспособности крови к образованию фибриновых сгустков, что приводит к
трудно поддающимся терапии кровотечениям.
Знание основ физиологии гемостаза позволяет клиницисту избрать
оптимальные варианты борьбы с заболеваниями, сопровождающимися тромбозами,
эмболиями, ДВС-синдромом и повышенной кровоточивостью.