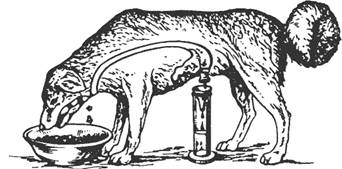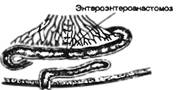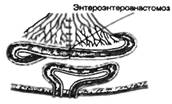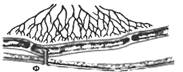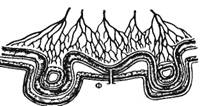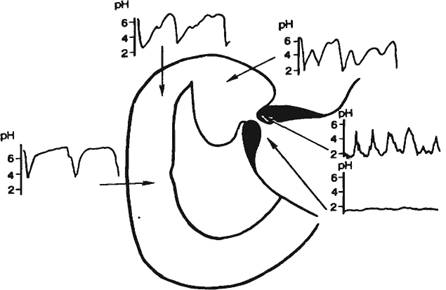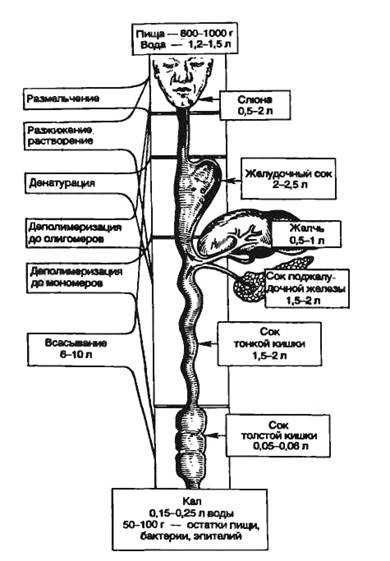
Рис. 9.1. Последовательность процессов в пищеварительном конвейере и выделение секретов в полость пищеварительного тракта.
Условием жизни является потребление энергетических и пластических веществ. Высшие животные в отличие от растений не могут их синтезировать из неорганических веществ, поэтому возникает необходимость их поступления из внешней среды. Длительное прекращение поступления или недостаточное введение в организм питательных веществ приводит к нарушению метаболизма и гомеостаза организма. Вместе с тем организм человека и высших животных не способен ассимилировать белки, жиры, углеводы и ряд других веществ пищи без их предварительной физико-химической обработки. Эту важную функцию выполняет система пищеварения.
9.1. ФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ ГОЛОДА И НАСЫЩЕНИЯ
В результате пищеварения и всасывания продуктов переваривания пищи поддерживается относительно постоянный уровень питательных веществ в организме. Лишение человека пищи влечет за собой состояние, называемое голодом. Прием пищи вызывает противоположное голоду чувство насыщения.
Голод. Как физиологическое состояние (в отличие от голодания как состояния длительной недостаточности питания, являющегося патологией) голод служит выражением потребности организма в питательных веществах, которых он был лишен на некоторое время, что привело к снижению их содержания в депо и циркулирующей крови.
Субъективным выражением голода служат неприятные ощущения жжения, «сосания под ложечкой», тошноты, иногда головокружения, головной боли и общей слабости. Внешним объективным проявлением голода является пищевое поведение, выражающееся в поиске и приеме пищи; оно направлено на устранение причин, вызывавших состояние голода. Субъективные и объективные проявления голода обусловлены возбуждением различных отделов ЦНС. Совокупность нервных элементов этих отделов И. П. Павлов назвал пищевым центром, функциями которого являются регуляция пищевого поведения и пищеварительных функций.
Пищевой центр — сложный гипоталамо-лимбико-ретикулокор-тикальный комплекс. Результаты экспериментов на животных свидетельствуют, что ведущим отделом являются латеральные ядра гипоталамуса. При их поражении наблюдается отказ от пищи (афагия), а при электрическом раздражении через вживленные в мозг электроды—повышенное потребление пищи (гиперфагия). Эту часть пищевого центра называют центром голода, или центром питания. Разрушение вентромедиальных ядер гипоталамуса приводит к гиперфагии, а их раздражение — к афагии. Считают, что в этих ядрах локализован центр насыщения. Между ним и центром голода установлены реципрокные отношения, т. е. если один центр возбужден, то другой заторможен. Описаны и более сложные отношения между этими ядрами.
Гипоталамические ядра представляют собой только часть (хотя и весьма важную) пищевого центра. Нарушение пищевого поведения происходит также и при поражении лимбической системы, ретикулярной формации и передних отделов коры больших полушарий.
Функциональное состояние гипоталамических ядер пищевого центра зависит от импульсов, поступающих с периферии от различных экстеро- и интероцепторов, состава и свойств притекающей к мозгу крови и находящейся в нем цереброспинальной жидкости. В зависимости от механизмов этих влияний предложено несколько теорий голода.
Теории голода. Одной из них признается, что ощущение голода и пищевое поведение вызываются импульсами от периодически сокращающегося свободного от пищи желудка. Эти сокращения желудка, повторяющиеся примерно через каждые l'/г ч и длящиеся 15—20 мин, назвали «голодными». При наполнении желудка пищей (и раздувании в нем резинового баллона) эти сокращения прекращаются и поступающие от желудка импульсы подавляют голод. Это так называемая локальная теория голода.
Однако имеются данные о том, что периодические сокращения желудка не совпадают с ощущениями голода у человека и проявлениями пищевого поведения у животных. Люди, у которых по соответствующим показаниям удален желудок, также ощущают голод. Периодические сокращения желудка есть у человека и некоторых плотоядных животных, а пищевое поведение характерно и для других видов. Собственно акт приема пищи, активация при этом секреции и моторики пищеварительного тракта имеют сигнальное значение и оказывают тормозное влияние на пищевой центр, вызывая так называемое первичное, или сенсорное, насыщение. Эти воздействия имеют значение в кратковременных изменениях состояния пищевого центра, а длительные изменения зависят от состояния метаболизма и отражающего его гомеостаза. Восстановление его после всасывания в кровь и депонирования пищевых веществ обеспечивает вторичное, обменное, или истинное, насыщение.
Насыщение. Это не только снятие чувства голода, но и ощущение удовольствия, полноты в желудке после приема пищи. Постепенно это ощущение угасает. В насыщении существенную роль играют психологические факторы, например привычка есть мало или много, в определенное время и т. д.
Состав крови голодных и накормленных человека и животных различен, что отражается на пищевом поведении последних: переливание голодному животному крови накормленного снижает у него пищевую мотивацию и количество принимаемой пищи. Имеются свидетельства различия свойств цереброспинальной жидкости накормленных и сытых животных.
Теории насыщения. В зависимости от вида веществ, с которыми связывают изменение состояния пищевого центра, предложено несколько теорий. Согласно глюкостатической теории, углеводный обмен влияет на аппетит через гипоталамические рецепторы, состояние которых изменяется в зависимости от различия в содержании глюкозы в артериальной и венозной крови. Аминацидостатическая теория отводит сигнальную роль аминокислотам крови, которые тормозят пищевой центр, особенно некоторые из них. Липостатическая теория связывает возбуждение пищевого центра с высвобождением липидов из жировых депо. Считают, что если глюкостатический механизм определяет кратковременные изменения состояния пищевого центра, то липостатический — долговременные его состояния.
Каждое из пищевых веществ используется в «метаболическом котле» организма, и его состояние контролируется пищевым центром по интегральным показателям. В роли сигналов об этом выступают теплообразование, компоненты цикла трикарбоновых кислот (Кребса), состояние энергетического баланса организма. Согласно термостатической теории, избыточное тепло, связанное с метаболическими процессами (и температурой окружающей среды), оказывает тормозное влияние на пищевой центр, является сигналом насыщения. Прием пищи увеличивает теплообразование (специфическое динамическое действие пищи) и участвует в формировании чувства насыщения. Метаболическая теория отводит сигнальную роль циркулирующим в крови ключевым компонентам цикла трикарбоновых кислот.
Роль таких метаболических сигналов и факторов, формирующих эти сигналы с помощью изменения метаболизма организма, деятельности пищеварительной системы, выполняют гормоны. К их числу относятся гормоны поджелудочной железы (инсулин, глюкагон, панкреатический полипептид), гипоталамо-гипофизар-ной системы (тиролиберин, кортиколиберин, нейротензин, соматотропин), щитовидной железы (тироксин, трийодтиронин, кальцитонин), гормоны пищеварительного тракта (арэнтерин, холецистокинин, гастрин, бомбезин), половые гормоны (эстрогены, андрогены), эндогенные и экзогенные опиаты (морфиноподобные вещества). Эти горомны изменяют функциональное состояние пищевого центра и как следствие — пищевое поведение.
В целом это зависит от интеграции экстеро- и интероцепторных нервных и различных активирующих и тормозных гуморальных влияний, а также влияний других центров мозга на пищевой центр.
Аппетит. В физиологии голода и насыщения применяется понятие «аппетит» (от лат. appetitus — стремление, желание) — ощущение, связанное со стремлением человека к определенной пище. Аппетит индивидуально вырабатывается и отражает не столько потребность в пище вообще, сколько потребность в связи со спецификой обмена веществ и дефицитом тех или иных компонентов пищи, индивидуальных и групповых привычек, особенностью национальной кухни и продуктов, которым отдается предпочтение. Избирательный аппетит — это влечение человека к определенному роду пищи, чаще тому, который содержит недостающие организму вещества.
У человека в норме выражены произвольная регуляция приема пищи и аппетита, управление количеством и качеством принимаемой пищи. Снижение и потерю аппетита вызывают многие факторы, в том числе те, которые отвлекают человека от еды, эмоции, в первую очередь отрицательные. Аппетит повышают острые и пряные приправы, закуски, предшествующие приему основных блюд, хорошая сервировка стола и др. Еда с аппетитом способствует эффективному пищеварению, расстраивает его прием пищи без аппетита. И. П. Павлов писал: «...врачи всех времен и стран до последнего времени считали своей существенной обязанностью, помимо борьбы с основными недугами, принимать специальные меры для восстановления аппетита».
Расстройства аппетита делят на три вида. Понижение его вплоть до полной потери называется анорексией (ап — отрицание, orexis — аппетит, греч.). Причинами ее является деятельность пищевого центра и пищеварительной системы, эндокринные, нейрогенные и психогенные расстройства, интоксикации.
Резкое повышение аппетита называют булимией (bu — бык, limes — голод, греч.). Она отмечается у некоторых больных с заболеваниями желудочно-кишечного тракта, при поражении головного мозга и его пищевого центра, некоторых эндокринных заболеваниях. Иногда булимия является результатом снижения чувства насыщения (акория), при этом наблюдается полифагия — прием чрезмерно большого количества пищи.
Еще один вид расстройств аппетита — его извращение, стремление принимать несъедобные вещества (мел, земля, уголь, керосин, бумага и т.д.). В одних случаях это результат выраженного специфического аппетита, в других — одно из психических расстройств и нарушения деятельности пищевого центра.
Физиология голода, аппетита, насыщения — раздел многих областей знаний. Безусловно, голод является одной из жизненно важных мотиваций, которая вызывает пищевое поведение, направленное на снятие этого состояния, отражающего сдвиги в гомеостазе организма. Восстановление гомеостаза и предоставление в распоряжение организма новых энергетических и пластических ресурсов происходит в результате приема и переработки пищи в процессе пищеварения.
9.2. СУЩНОСТЬ ПИЩЕВАРЕНИЯ. КОНВЕЙЕРНЫЙ ПРИНЦИП ОРГАНИЗАЦИИ ПИЩЕВАРЕНИЯ
9.2.1. Пищеварение и его значение
Пищеварение — сложный физиологический и биохимический процесс, в ходе которого принятая пища в пищеварительном тракте подвергается физическим и химическим изменениям. В результате этого компоненты пищи должны сохранить свою пластическую и энергетическую ценность; приобрести свойства, благодаря которым они могут быть усвоенными организмом и включенными в его нормальный обмен веществ; утратить видовую специфичность (при сохранении которой компоненты пищи не усваиваются и как чужеродные вещества, вызывающие защитные реакции организма, могут быть причиной тяжелых патологических явлений).
Физические изменения пищи состоят в ее размельчении, набухании, растворении, химические — в последовательной деградации питательных веществ в результате действия на них компонентов пищеварительных соков, выделяемых в полость пищеварительного тракта его железами. Важнейшая роль в этом принадлежит гидролитическим ферментам секретов пищеварительных желез и исчерченной каемки тонкой кишки.
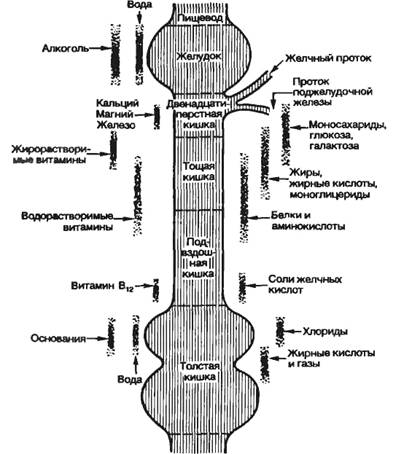
Названные процессы идут в определенной последовательности, «наслаиваясь» по отделам пищеварительного тракта (рис. 9.1). Продвижение пищевого содержимого в дистальном направлении, его задержка на различное время в том или ином отделе пищеварительного тракта, смешивание пищевых веществ с пищеварительным секретами обеспечиваются его гладкомышечным аппаратом, т. е. моторный аппарат пищеварительного тракта распределяет пищеварение во времени и пространстве и в большой мере влияет на его интенсивность. В результате деполимеризации питательных веществ образуются продукты, в основном мономеры, которые всасываются из кишечника в кровь и лимфу, транспортируются к тканям организма и включаются в его метаболизм. Вода, минеральные соли и некоторые органические компоненты пищи (в том числе витамины) всасываются в кровь неизмененными.
В зависимости от происхождения гидролитических ферментов пищеварение делят на три типа (А. М. Уголев): собственное, симбионтное и аутолитическое.
|
|
|
Рис. 9.1. Последовательность процессов в пищеварительном конвейере и выделение секретов в полость пищеварительного тракта. |
Собственное пищеварение осуществляется ферментами, синтезированными данным макроорганизмом, его железами, эпителиальными клетками — ферментами слюны, желудочного и поджелудочного соков, эпителия тонкой кишки.
Симбионтное пищеварение — гидролиз питательных веществ за счет ферментов, синтезированных симбионтами макроорганизма — бактериями и простейшими пищеварительного тракта. Симбионтное пищеварение у человека осуществляется в толстой кишке. У человека клетчатка пищи по типу собственного пищеварения из-за отсутствия соответствующего фермента в секретах желез не гидролизуется (в этом заключается определенный физиологический смысл — сохранение пищевых волокон, играющих важную роль в кишечном пищеварении), поэтому переваривание ее ферментами симбионтов в толстой кишке является важным процессом. В результате симбионтного пищеварения образуются вторичные пищевые вещества в отличие от первичных, образующихся в результате собственного пищеварения. У человека в условиях развитого собственного пищеварения его роль в общем пищеварительном процессе относительно невелика. Аутолитическое пищеварение осуществляется за счет экзогенных гидролаз, которые вводятся в организм в составе принимаемой пищи. Роль данного пищеварения существенна при недостаточно развитом собственном пищеварении. У новорожденных собственное пищеварение еще не развито, поэтому возможно его сочетание с аутолитическим пищеварением, т. е. питательные вещества грудного молока перевариваются ферментами, поступающими в пищеварительный тракт младенца в составе грудного молока.
В зависимости от локализации процесса гидролиза питательных веществ пищеварение делят на несколько типов. Прежде всего на внутри- и внеклеточное.
Внутриклеточное пищеварение состоит в том, что транспортированные в клетку путем фагоцитоза и пиноцитоза (эндоцитоза) вещества гидролизуются клеточными (лизосомальными) ферментами либо в цитозоле, либо в пищеварительной вакуоли. Эндоцитозу отводится значительная роль в кишечном пищеварении в период раннего постнатального развития млекопитающих. В последнее время получены электронно-микроскопические данные о достаточно высокой, но качественно трансформированной эндоцитозной активности энтероцитов взрослых млекопитающих (И. А. Морозов).
Внеклеточное пищеварение делят (А. М. Уголев) на дистантное и контактное, пристеночное, или мембранное. Дистантное пищеварение совершается в среде, удаленной от места продукции гидролаз. Так осуществляется действие на питательные вещества в полости пищеварительного тракта ферментов слюны, желудочного сока и сока поджелудочной железы. Такое пищеварение в специальных полостях называется полостным. Эффективность полостного пищеварения определяется активностью ферментов секретов пищеварительных желез в соответствующих отделах пищеварительного тракта.
Пристеночное, контактное, или мембранное, пищеварение открыто в 50-х годах текущего столетия А. М. Уголевым. Такое пищеварение происходит в тонкой кишке на колоссальной поверхности, образованной складками, ворсинками и микроворсинками ее слизистой оболочки. Гидролиз происходит с помощью ферментов, «встроенных» в мембраны микроворсинок.
Богаты ферментами слизь, выделяемая слизистой оболочкой тонкой кишки (Ю. М. Гальперин и др.), и зона исчерченной каемки, образованная микроворсинками и мукополисахаридными нитями — гликокаликсом. В слизи и гликокаликсе находятся ферменты поджелудочной железы, перешедшие из полости тонкой кишки, и собственно кишечные ферменты, образующиеся в результате непрерывных процессов кишечной секреции и отторжения энтероцицитов.
Следовательно, пристеночное пищеварение в широком его понимании совершается в слое слизи, зоне гликокаликса и на поверхности микроворсинок с участием большого количества ферментов кишки и поджелудочной железы.
В настоящее время процесс пищеварения рассматривают как трехэтапный: полостное пищеварение- пристеночное пищеварение- всасывание. Полостное пищеварение заключается в начальном гидролизе полимеров до стадии олигомеров, пристеночное обеспечивает дальнейшую ферментативную деполимеризацию олигомеров в основном до стадии мономеров, которые затем всасываются.
9.2.3. Конвейерный принцип организации пищеварения
И. П. Павлов сравнивал деятельность пищеварительного тракта с конвейерным химическим производством. Этот «конвейер» представляется в виде последовательной цепи деградации пищи и ее питательных веществ. Пищеварительный конвейер заключается в преемственности следующих процессов:
1) органных: пищеварение в полости рта- желудочное пищеварение - кишечное пищеварение;
2) физических и химических: размельчение, увлажнение, набухание, растворение пищи; денатурация белков; гидролиз полимеров до стадии различных олигомеров, затем мономеров; их транспорт из пищеварительного тракта в кровь и лимфу;
3) полостного и пристеночного пищеварения от центральной части пищевого комка в желудке к его примукозальному слою;от вершины кишечной ворсинки к ее основанию; от полостного гидролиза питательных веществ в тонкой кишке к продолжению его в зоне примукозальной слизи, затем в зоне гликокаликса и наконец на мембранах энтероцитов;
4) гидролиза на апикальных мембранах энтероцитов и транспорта в энтероцит образовавшихся мономеров, а из него — в интерстициальную ткань и затем в кровь и лимфу;
5) ферментативной деполимеризации питательных веществ.
При этом в каждом проксимальнее расположенном отделе осуществляются
процессы, повышающие эффективность их в следующем, дистальнее расположенном
отделе.
Интеграция, правильная последовательность работы элементов пищеварительного конвейера во времени и пространстве обеспечиваются регуляторными процессами различного уровня.
Ферментативная активность свойственна каждому отделу пищеварительного тракта и максимальна при определенном значении рН среды. Например, в желудке пищеварительный процесс осуществляется в кислой среде. Переходящее в двенадцатиперстную кишку кислое содержимое нейтрализуется, и кишечное пищеварение происходит в нейтральной и слабоосновной среде, созданной выделяющимися в кишку секретами — желчью, соками поджелудочной железы и кишечным, которые инактивируют желудочные пепсины. Кишечное пищеварение происходит в нейтральной и слабоосновной среде сначала по типу полостного, а затем пристеночного пищеварения, завершающегося всасыванием продуктов гидролиза (нутриентов). В нормальных условиях основной гидролиз пищевых веществ завершается в проксимальном отделе тонкой кишки, а дистальный ее отдел является резервным, включающимся в пищеварительный процесс с целью его компенсации при функциональной недостаточности проксимального отдела кишечника.
Переваривание пищевых веществ в пищеварительном тракте. Деградация пищевых веществ по типу полостного и пристеночного пищеварения осуществляется гидролитическими ферментами. Каждый из них имеет выраженную в той или иной мере субстратную специфичность. Набор ферментов в составе секретов пищеварительных желез и исчерченной каемки тонкой кишки имеет видовую и индивидуальную особенности, адаптирован к перевариванию той пищи, которая характерна для данного вида животного, и тем питательным веществам, которые преобладают в рационе.
9.3. ПИЩЕВАРИТЕЛЬНЫЕ ФУНКЦИИ ПИЩЕВАРИТЕЛЬНОГО ТРАКТА
Пищеварительный тракт (желудочно-кишечный тракт) — часть пищеварительной системы, имеющая трубчатое строение и включающая пищевод, желудок, толстую и тонкую кишку, в которых происходят механическая и химическая обработка пищи и всасывание продуктов гидролиза.
9.3.1. Секреция пищеварительных желез
Секреция — внутриклеточный процесс образования из веществ, поступивших в клетку, специфического продукта (секрета) определенного функционального назначения и выделение его из железистой клетки. Секреты поступают через систему секреторных ходов и протоков в полость пищеварительного тракта.
Секреция пищеварительных желез обеспечивает доставку в полость пищеварительного тракта секретов, ингредиенты которых гидролизуют питательные вещества (секреция гидролитических ферментов и их активаторов), оптимизируют условия для этого (по рН и другим параметрам — секреция электролитов) и состояние гидролизуемого субстрата (эмульгирование липидов солями желчных кислот, денатурация белков соляной кислотой), выполняют защитную роль (слизь, бактерицидные вещества, иммуноглобулины). .
Секреция пищеварительных желез контролируется нервными, гуморальными и паракринными механизмами. Эффект этих влияний — возбуждение, торможение, модуляция секреции гландулоцитов — зависит от вида эфферентных нервов и их медиаторов, гормонов и других физиологически активных веществ, гландулоцитов, мембранных рецепторов на них, механизма действия этих веществ на внутриклеточные процессы. Секреция желез находится в прямой зависимости от уровня их кровоснабжения, которое в свою очередь определяется секреторной активностью желез, образованием в них метаболитов — вазодилататоров, влиянием стимуляторов секреции как вазодилататоров. Количество секрета железы зависит от числа одновременно секретирующих в ней гландулоцитов. Каждая железа состоит из гландулоцитов, вырабатывающих разные компоненты секрета, и имеет существенные особенности регуляции. Это обеспечивает широкое варьирование состава и свойств выделяемого железой секрета. Он изменяется также по мере продвижения по протоковой системе желез, где некоторые компоненты секрета всасываются, другие выделяются в проток его гландулоцитами. Изменения количества и качества секрета адаптированы к виду принятой пищи, составу и свойствам содержимого пищеварительного тракта.
Для пищеварительных желез основными стимулирующими секрецию нервными волокнами являются парасимпатические холинергические аксоны постганглионарных нейронов. Парасимпатическая денервация желез вызывает разной длительности (на несколько дней и недель) гиперсекрецию желез (особенно слюнных, в меньшей мере желудочных) — паралитическую секрецию, в основе которой лежит несколько механизмов (см. раздел 9.6.3).
Симпатические нейроны тормозят стимулированную секрецию и оказывают на железы трофические влияния, усиливая синтез компонентов секрета. Эффекты зависят от вида мембранных рецепторов — α- и β -адренорецепторов, через которые они реализуются.
В роли стимуляторов, ингибиторов и модуляторов секреции желез выступают многие гастроинтестинальные регуляторные пептиды.
В естественных условиях количество, состав и динамика секреции пищеварительных желез определяются соотношением одновременно и последовательно действующих регуляторных механизмов.
9.3.2. Моторная функция пищеварительного тракта
Моторная, или двигательная, функция осуществляется на всех этапах процесса пищеварения. В пищеварительном тракте происходят произвольные и непроизвольные, макро- и микромоторные явления. Прием, механическая переработка пищи в ходе жевания, глотание, задержка в желудке и эвакуация его содержимого в кишечник, сокращения и расслабления желчного пузыря, перемешивание и передвижение кишечного содержимого (химуса), перераспределение давления в отделах тонкой кишки, перемешивание пристеночного слоя химуса, переход химуса из тонкой кишки в толстую, сокращение и расслабление сфинктеров, движения толстой кишки, необходимые для формирования кала и дефекации,— основные моторные процессы, обеспечивающие процесс пищеварения в различных отделах пищеварительного тракта.
Изменение тонуса и перистальтики выводных протоков пище варительных желез, состояние их сфинктеров обеспечивают выведение пищеварительных секретов. К моторике также относятся движения ворсинок и микроворсинок.
Гладкие мышцы пищеварительного тракта образованы гладкими мышечными клетками (миоциты), обладающими рядом специфических физиологических свойств (см. раздел 2.4.1.7). Миоциты плотно упакованы в пучки и соединены нексусами. Пучок считается функциональной единицей гладкой мышцы. Пучок иннервируется нервными терминалями, он также получает мелкую артериолу. Нейромедиаторы и физиологически активные вещества, вышедшие из крови в интерстициальную жидкость пучка, оказывают на его миоциты возбуждающие и тормозные влияния.
Гладкие мышцы пищеварительного тракта относятся к группе унитарных и обладают способностью спонтанного ритмического возбуждения и свойствами синцития. Растяжение гладких мышц вызывает деполяризацию их мембран и мышечное сокращение. Вегетативные нервы, гормоны и парагормоны изменяют частоту и силу этих сокращений в широких пределах. На протяжении пищеварительного тракта имеется несколько водителей ритма его сокращений. Эти водители ритма особенно чувствительны к физиологически активным веществам и получают обильную иннервацию.
Сложность движений пищеварительного тракта обеспечивается наличием в нем слоев и пучков гладких мышц, идущих в разных направлениях, при расслаблении или сокращении которых уменьшается или увеличивается тонус кишки и изменяется просвет пищеварительного канала. Волна сокращений и расслабления круговых мышц продвигается вдоль пищеварительного канала, создавая его перистальтические сокращения. Согласование сокращений различных мышечных пучков осуществляется посредством периферической интрамуральной нервной системы.
В пищеварительном тракте около 35 сфинктеров (жомов) — специальных замыкательных аппаратов, состоящих из скопления преимущественно циркулярно расположенных мышечных пучков, а также мышечных пучков спирального и продольного направлений. Сокращение циркулярно расположенных мышечных пучков обеспечивает смыкание и уменьшение просвета сфинктера, сокращение спирально и продольно расположенных пучков увеличивает просвет сфинктера. Сфинктеры выполняют роль клапанов, обеспечивающих движение пищевого содержимого в каудальном направлении, одноправленное движение пищеварительных секретов, разобщение отделов пищеварительного тракта, где пищеварение происходит на характерных для них этапах.
В координации моторики пищеварительного тракта велика роль миогенных механизмов, периферической (интра- и экстрамуральной) и центральной нервной системы. Последняя имеет важное значение в пусковых влияниях на органы пищеварения, в изменении их реактивности, интеграции моторной и секреторной функций пищеварительного тракта, его адаптации к виду принятой пищи. Парасимпатические влияния преимущественно повышают моторную активность пищеварительного тракта, но в составе блуждающих нервов имеются возбуждающие и тормозящие моторику нервные волокна. Симпатические влияния заключаются в основном в снижении моторной активности. Нервные, гормональные и пара-гормональные влияния создают сочетанные органные и межорганные внутрисистемные эффекты. Так, желчевыделение осуществляется сокращениями желчного пузыря при открытом сфинктере печеночно-поджелудочной ампулы (сфинктер Одди); желудочная эвакуация — при сокращении антральной части желудка, но расслабленном сфинктере привратника (пилорический сфинктер).
Всасывание — процесс транспорта компонентов пищи из полости пищеварительного тракта во внутреннюю среду, кровь и лимфу организма. Всосавшиеся вещества разносятся по организму и включаются в обмен веществ тканей. В полости рта химическая обработка пищи сводится к частичному гидролизу углеводов амилазой слюны, при котором крахмал расщепляется на декстрины, мальтоолигосахариды и мальтозу. Кроме того, время пребывания пищи в полости рта незначительно, поэтому всасывания здесь практически не происходит. Однако известно, что некоторые фармакологические вещества всасываются быстро, и это находит применение как способ введения лекарственных веществ.
В желудке всасывается небольшое количество аминокислот, глюкозы, несколько
больше воды и растворенных в ней минеральных солей, значительно всасывание
растворов алкоголя. Всасывание питательных веществ, воды, электролитов
осуществляется в основном в тонкой кишке и сопряжено с гидролизом
питательных веществ. Всасывание зависит от величины поверхности, на которой
оно осуществляется. Особенно велика поверхность всасывания в тонкой кишке.
У человека поверхность слизистой оболочки тонкой кишки увеличена в 300—500 раз
за счет складок, ворсинок и микроворсинок. На 1 мм* слизистой оболочки
кишки приходится 30—40 ворсинок, а каждый энтероцит имеет 1700—4000
микроворсинок. На
У взрослого человека число всасывающих кишечных клеток составляет 10'°, а соматических клеток — 10'°. Из этого следует, что одна кишечная клетка обеспечивает питательными веществами около 100 000 других клеток организма человека. Это предполагает высокую активность энтероцитов в гидролизе и всасывании питательных веществ. Микроворсинки покрыты слоем гликокаликса,образующего из мукополисахаридных нитей на апикальной поверхности слой толщиной до 0,1 мкм. Нити связаны между собой кальциевыми мостиками,что обуславливает формирование особой сети. Она обладает свойствами молекулярного сита, разделющего молекулы по их величине и заряду. Сеть имеет отрицательный заряд и гидрофильна, что придает направленный и селективный характер транспорту через нее низкомолекулярных веществ к мембране микроворсинок, препятствует транспорту через нее высокомолекулярных веществ и ксенобиотиков. Гликокаликс удерживает на поверхности эпителия кишечную слизь, которая вместе с гликокаликсом адсорбирует из полости кишки гидролитические ферменты, продолжающие полостной гидролиз питательных веществ, продукты которого переводятся на мембранные системы микроворсинок. На них завершается гидролиз питательных веществ по типу мембранного пищеварения с помощью кишечных ферментов с образованием в основном мономеров, которые всасываются.
Всасывание различных веществ осуществляется разными механизмами.
Всасывание макромолекул и их агрегатов происходит путем фагоцитоза и пиноцитоза. Эти механизмы относятся к эндоцитозу. С эндоцитозом связано внутриклеточное пищеварение, однако ряд веществ, попав в клетку путем эндоцитоза, транспортируется в везикуле через клетку и выделяется из нее путем экзоцитоза в межклеточное пространство. Такой транспорт веществ назван трансцитозом. Он, видимо, из-за небольшого объема не имеет существенного значения во всасывании питательных веществ, но важен в переносе иммуноглобулинов, витаминов, ферментов и т. д. из кишечника в кровь. У новорожденных трансцитоз важен в транспорте белков грудного молока.
Некоторое количество веществ может транспортироваться по межклеточным пространствам. Такой транспорт называется персорбцией. С помощью персорбции переносятся часть воды и электролитов, а также другие вещества, в том числе белки (антитела, аллергены, ферменты и т. п.) и даже бактерии.
В процессе всасывания микромолекул — основных продуктов гидролиза питательных веществ в пищеварительном тракте, а также электролитов участвует три вида транспортных механизмов: пассивный транспорт, облегченная диффузия и активный транспорт. Пассивный транспорт включает в себя диффузию, осмос и фильтрацию. Облегченная диффузия осуществляется с помощью особых мембранных переносчиков и не требует затраты энергии. Активный транспорт — перенос веществ через мембраны против электрохимического или концентрационного градиента с затратой энергии и при участии специальных транспортных систем (мембранные транспортные каналы, мобильные переносчики, конформационные переносчики). Мембраны имеют транспортеры многих типов. Эти молекулярные устройства переносят один или несколько типов веществ. Часто транспорт одного вещества сопряжен с движением другого вещества, перемещение которого по градиенту концентрации служит источником энергии для сопрягаемого транспорта. Чаще всего в такой роли используется электрохимический градиент Na+. Натрийзависимым процессом в тонкой кишке является всасывание глюкозы, галактозы, свободных аминокислот, дипептидов и трипептидов, солей желчных кислот, билирубина и ряда других веществ. Натрийзависимый транспорт осуществляется и через специальные каналы, и посредством мобильных переносчиков. Натрийзависимые транспортеры расположены на апикальных мембранах, а натриевые насосы — на базолатеральных мембранах энтероцитов. В тонкой кишке существует и натрий-независимый транспорт многих мономеров пищевых веществ. Транспортные механизмы клеток связаны с деятельностью ионных насосов, которые используют энергию АТФ с помощью Na+, К+-АТФазы. Она обеспечивает градиент концентраций натрия и калия между вне- и внутриклеточной жидкостями и, следовательно, участвует в обеспечении энергией натрийзависимого транспорта (и мембранных потенциалов). Na+, К+-АТФаза локализована в базолатеральной мембране. Последующее откачивание ионов Na+ из клеток через базолатеральную мембрану (что создает градиент концентрации натрия на апикальной мембране) связано с затратой энергии и участием Na+, К+-АТФаз этих мембран. Транспорт мономеров (аминокислот и глюкозы), образовавшихся в результате мембранного гидролиза димеров на апикальной мембране кишечных эпителиоцитов, не требует участия ионов Na+ и обеспечивается энергией ферментно-транспортного комплекса. Мономер передается с фермента этого комплекса в транспортную систему без предварительного перевода в премембранную водную фазу.
Скорость всасывания зависит от свойств кишечного содержимого. Так, при прочих равных условиях всасывание идет быстрее при нейтральной реакции этого содержимого, чем при кислой и щелочной; из изотонической среды всасывание электролитов и питательных веществ происходит быстрее, чем из гипо- и гипертонической среды. Активное создание в пристеночной зоне тонкой кишки с помощью двустороннего транспорта веществ слоя с относительно постоянными физико-химическими свойствами является оптимальным для сопряженного гидролиза и всасывания питательных веществ.
Повышение внутрикишечного давления увеличивает скорость всасывания из тонкой кишки раствора поваренной соли. Это указывает на значение фильтрации во всасывании и роль кишечной моторики в этом процессе. Моторика тонкой кишки обеспечивает перемешивание пристеночного слоя химуса, что важно для гидролиза и всасывания его продуктов. Доказано преимущественное всасывание разных веществ в различных отделах тонкой кишки. Допускается возможность специализации разных групп энтероцитов на преимущественной резорбции тех или иных пищевых веществ.
Большое значение для всасывания имеют движения ворсинок слизистой оболочки тонкой кишки и микроворсинок энтероцитов. Сокращениями ворсинок лимфа с всосавшимися в нее веществами выдавливается из сжимающейся полости лимфатических .сосудов. Наличие в них клапанов препятствует возврату лимфы в сосуд при последующем расслаблении ворсинки и создает присасывающее действие центрального лимфатического сосуда. Сокращения микроворсинок усиливают эндоцитоз и, возможно, являются одним из его механизмов.
Натощак ворсинки сокращаются редко и слабо, при наличии в кишке химуса сокращения ворсинок усилены и учащены (до 6 в 1 мин у собаки). Механические раздражения основания ворсинок вызывают усиление их сокращений, тот же эффект наблюдается под влиянием химических компонентов пищи, особенно продуктов ее гидролиза (пептиды, некоторые аминокислоты, глюкоза и экстрактивные вещества пищи). В реализации этих воздействий определенная роль отводится интрамуральной нервной системе (подслизистое, или мейснеровское, сплетение).
Кровь сытых животных, перелитая голодным, вызывает у них усиление движения ворсинок. Считают, что при действии кислого желудочного содержимого на тонкую кишку в ней образуется гормон вилликинин, который через кровоток стимулирует движения ворсинок. В очищенном виде вилликинин не выделен. Скорость всасывания из тонкой кишки в большой мере зависит от уровня ее кровоснабжения. В свою очередь оно увеличивается при наличии в тонкой кишке продуктов, подлежащих всасыванию.
Всасывание питательных веществ в толстой кишке незначительно, так как при нормальном пищеварении большая часть их уже всосалась в тонкой кишке. В толстой кишке всасывается большое количество воды, в небольшом количестве могут всасываться глюкоза, аминокислоты и некоторые другие вещества. На этом основано применение так называемых питательных клизм, т. е. введение легкоусвояемых питательных веществ в прямую кишку.
Всасывание различных веществ имеет существенные различия, которые рассматриваются в разделе 9.6.6.
9.3.4. Методы изучения пищеварительных функций
9.3.4.1. Экспериментальные методы
Острые опыты. Острые опыты на наркотизированных животных продолжают применять для решения аналитических задач. Для этих же целей используют опыты на изолированных in vitro органах, тканях и клетках. Например, мембранное пищеварение было открыто и детально исследовано в острых опытах на изолированных отрезках тонкой кишки крыс, однако перенос результатов таких опытов на деятельность целого организма ограничен.
Методы хронического эксперимента. Принцип хронического эксперимента заключается в хирургической (оперативной) подготовке животных, в ходе которой накладывают фистулу (отверстие, снабженное специальной трубкой, выходящей наружу) того или иного отдела пищеварительного тракта или выводных протоков пищеварительных желез. Опыты ставят на выздоровевщих после операции животных.
|
|
|
Рис. 9.2. Эзофаготомированная собака с фистулой желудка. Опыт «мнимого кормления». |
В. А. Басов (1842) успешно произвел операцию наложения фистулы желудка у собак. При дальнейшем усовершенствовании этой операции в желудочном свище фиксировали трубку, которую вне опыта закрывали пробкой. Открыв ее, можно было получать содержимое желудка.
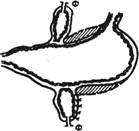
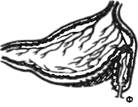
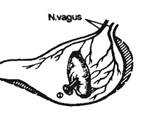
В лаборатории И. П. Павлова у таких собак была выполнена операция эзофаготомии (перерезка пищевода). После заживления раны производили «мнимое кормление» собаки: она ела, но пища выпадала из отверстия пищевода, а из открытой желудочной фистулы изливался сок (рис. 9.2). Сок в чистом виде получали у собак с изолированными выкроенными в хирургических операциях из различных частей желудка желудочками (рис. 9.3). Желудочек, выкроенный по методу Павлова, в отличие от желудочка Гейденгайна имеет сохраненную вагусную иннервацию и более полно отражает секрецию в большом желудке, где идет пищеварительный процесс. Применяют методы хирургической изоляции петли тонкой кишки с выведением в кожную рану одного дистального (операция Тири) или двух (операция Тири—Веяла) ее концов (рис. 9.4), из которых собирают кишечный сок или куда вводят растворы для изучения их всасывания.
Широкое распространение получили операции выведения наружу и вживления в кожную рану выводных протоков слюнных и поджелудочной желез, желчного выводного протока. Разработаны методы, предотвращающие потерю пищеварительных секретов вне экспериментов.
Фистульная методика позволяет в любое время наблюдать за функцией органа, который имеет нормальные кровоснабжение и иннервацию. Из фистулы собирают чистые пищеварительные соки, изучают их состав и свойства натощак, после кромления животных или иной стимуляции секреции. На фистульных животных изучают моторную и секреторную функции органов пищеварения, процессы гидролиза и всасывания питательных веществ в различных отделах
|
1 |
2 |
|
3 |
4 |
|
5 |
6 |
|
7 |
8 |
|
Рис. 9.3. Варианты операций по созданию изолированного желудочка (схема). 1 — неоперированный желудок с блуждающими нервами, идущими от пищевода по большой и малой кривизне; 2 — желудочек по И. П. Павлову; 3 — по Гейденгайну; 4 — изолированные желудочки большой и малой кривизны с «мостиками» со стороны кардан; 5 — то же с «мостиками» со стороны пилорической части желудочка; б — изолированный желудочек передней стенки желудка; 7 — изолированный пилорический желудочек и двенадцатиперстная кишка; 8 — то же, что и 7, но с сохранением серозно-мышечного слоя и нервов со стороны фундальной части желудка и двенадцатиперстной кишки, ф — фистульная трубка из металла или пластмассы. | |
пищеварительного тракта на практически здоровых животных в почти естественных условиях хронических экспериментов. В исследованиях И. П. Павлова, принесших ему широкую славу и Нобелевскую премию (1904), в хронических опытах были получены новые данные, и, как сказано в Нобелевском дипломе, И. П. Павлов «пересоздал физиологию пищеварения».
9.3.4.2. Исследование пищеварительных функций у человека
С исследовательской целью фистулы человеку не накладывают. Иногда фистулы образуются при ранении, иной патологии, их делают с целью сохранения жизни человека, например для введения пищи в желудок при непроходимости пищевода. Основные методы исследования пищеварительных функций у человека ориентированы на их безвредность и безболезненность. Эти методы используют в функциональной диагностике здорового и больного человека.
|
|
||
|
Рис.9.5. Собирание слюны у человека с помощью капсулы Лешли-Красногорского. 1-трубка для отсасывания воздуха из внешней камеры капсулы;2-трубка для слюны из внутренней камеры капсулы. | ||
|
1 |
|
2 |
|
3 |
|
4 |
|
Рис. 9.4. Варианты изолированной петли тонкой кишки (схема). 1-изолированная петля в выведенным в кожную рану дистальным ее концом; 2-петля с изолированной полостью(серозно-мышечный слой сохранен);4-в кожные лоскуты выведены два участка тонкой кишки, пережатие их изолирует во время опыта полость кишечной петли с фистульной трубкой (Ф). |
Исследование процессов секреции.
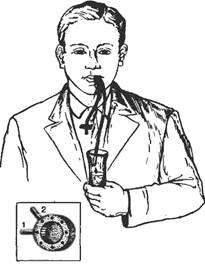
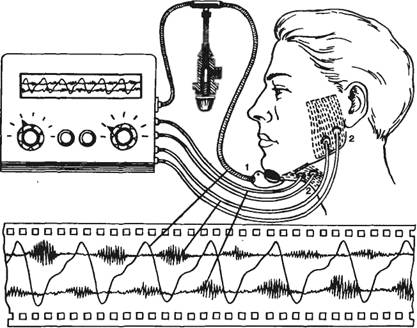
Для изучения слюноотделения слюну получают при сплевывании после полоскания рта, но получаемая при этом ротовая жидкость является смесью слюны разных желез, остатков пищи и других компонентов полости рта; кроме того, нельзя точно определить ее объем. Чистую слюну крупных слюнных желез получают путем катетеризации их протоков и с помощью капсул Лешли—Красногорского, фиксируемых к слизистой оболочке рта над протоками околоушных, поднижнечелюстной и подъязычной слюнных желез (у них проток открывается единым сосочком). Человек с капсулой во рту (рис. 9.5) может жевать пищу, что вызывает саливацию. Применяют и другие ее стимуляторы. Учитывают объем выделившейся за определенное время слюны, определяют ее состав и свойства (вязкость, рН, содержание электролитов, ферментов, муцина).
Для изучения секреторной деятельности желез желудка, поджелудочной железы, тонкой кишки, желчевыделения у человека используют зондовые и беззондовые методы. При зондовых исследованиях испытуемый проглатывает (или ее вводят через нос) эластичную трубку, которая проводится в желудок, двенадцатиперстную или тощую кишку. Существуют двухканальные зонды для одновременного получения содержимого желудка и двенадцатиперстной кишки, которое можно отсасывать как натощак, как и после стимуляции пищеварительных желез различными методами (прием пробного завтрака, различных фармакологических стимуляторов и т. д.).
Применение эндоскопических управляемых зондов позволило вводить тонкий катетер в проток поджелудочной железы и получать ее секрет без примеси к нему других секретов, что неизбежно при аспирации содержимого двенадцатиперстной кишки.
Зондовые методы позволяют определять объем секрета и содержание различных его компонентов: электролитов, ферментов, а также рН и др. Стимуляторы секреции вводят в пищеварительный тракт или парентерально. Знание механизмов их действия позволяет определить место, характер и причины нарушения секреции.
Существуют методы зондирования, с помощью которых возможно определение ряда параметров непосредственно в полости пищеварительного тракта, наблюдения за их динамикой в содержимом желудка или кишечника. Для этого зонды снабжают соответствующими датчиками (например, датчиками рН, давления, электродами для отведения регистрируемых потенциалов и др.). Методы эндоскопического исследования желудка и кишечника, кроме визуального контроля за состоянием слизистой оболочки, позволяют брать ее кусочки для последующего морфологического и биохимического исследования.
Наконец, существуют зонды, с помощью которых полость желудка или кишки перфузируют растворами разного состава. Так, перфузируя отрезок кишки раствором какого-либо вещества, например крахмала, можно по разности его концентрации во вводимом и аспирируемом растворах определить переваривание крахмала и оценить соответствующую ферментативную активность исследуемого отрезка кишки.
Применение зондовых методов в ряде случаев противопоказано, поэтому разрабатываются и беззондовые, основанные на разных принципах методы исследования секреции пищеварительных желез. В одних методах учитывают содержание в крови и выделение с мочой веществ, освободившихся из принятых препаратов под действием на них пищеварительных секретов. Например, если кислотность желудочного сока нормальная, то индикатор быстро появляется в крови и моче, если кислотность низкая или нулевая, то в исследуемых жидкостях индикатор отсутствует или появляется с большим опозданием.
В другой группе беззондовых методов функциональное состояние пищеварительных желез оценивают по активности их ферментов в крови и моче: она при прочих равных условиях тем выше, чем большее число гландулоцитов тех или иных желез синтезирует данные ферменты, которые покидают железы не только в составе секретов, но частично транспортируются в лимфу и кровь, откуда выводятся в составе мочи (и других экскретов).
Косвенно оценить полноценность секреции пищеварительных желез можно по наличию в кале негидролизованных компонентов принятой пищи, а также определяя активность в кале ферментов поджелудочной железы и тонкой кишки.
С развитием радиотелеметрии появилась возможность сконструировать приборы для эндорадиозондирования пищеварительного тракта. Проглоченная радиокапсула, передвигаясь по нему, может в виде радиосигналов передавать информацию о ряде параметров его содержимого, в том числе о pН.
|
|
|
Рис. 9.6. Регистрация функции жевательной мускулатуры методом миоэлектрома- стикациографии (по И. С. Рубину). 1 — мастикациограф; 2 — электроды для отведения биопотенциалов жевательных мышц. |
Исследование моторной функции. Методика исследования акта жевания (мастикациография — графическая регистрация жевательных движений нижней челюсти) характеризует его длительность и длительность составляющих жевание фаз, координированность акта. Иногда регистрация движений нижней челюсти в этом методе сочетается с электромиографией жевательных мышц (рис. 9.6). Методом гнатодинамометрии оценивают давление, которое развивается жевательными мышцами на разных парах зубов при смыкании челюстей. Результативность жевания может быть учтена по размерам пищевых частиц в составе пищевого комка, подготовленного к глотанию.
Глотание исследуют рентгенографически и рентгенокинематографически, баллонографически с помощью специальных зондов. Нашли применение также многоканальные зонды, с помощью которых регистрируют давление в пищеводе при продвижении по нему перистальтическими волнами заглатываемого пищевого комка. Применяют также эзофагоскопию и аускультацию глотательных шумов.
Моторную активность желудка и кишечника, как и секрецию, изучают зондовыми и беззондовыми методами. Зондовые методы предполагают использование зондов с резиновыми баллончиками или свободных на конце зондов, наполненных изотоническим раствором натрия хлорида, через который передает ся давление в полости желудка и тонкой кишки на воспринимающие и регистрирующие устройства. Используют многоканальные зонды, позволяющие регистрировать давление в нескольких отделах желудка и тонкой кишки.
Сконструированы комбинированные зонды, позволяющие одновременно регистрировать давление в полостях желудка и кишечника, рН, биопотенциалы, аспирировать содержимое, вводить тестовые стимуляторы секреции и моторики.
Беззондовым методом изучения моторной активности пищеварительного тракта является радиотелеметрический, при котором используется радиокапсула (радиопилюля) с датчиком давления. Моторную активность желудка можно оценить электрографически, отводя медленные потенциалы гладких мышц сокращающегося желудка с передней брюшной стенки. Этот метод иногда используют для регистрации моторной активности тонкой и толстой кишки.
Широкое распространение в клинике получили методы рентгенологического изучения моторики пищевода, желудка, кишечника, желчного пузыря и желчных путей, заполненных рентгеноконтрастным веществом.
Моторную активность органов пищеварения оценивают также по скорости и динамике эвакуации из желудка его содержимого в кишечник и продвижению содержимого по нему. Для этого используют рентгенологические и радиологические методы, в том числе радиоизотопное сканирование. В этих методах к принимаемой пище добавляют безвредное количество изотопа с коротким периодом распада и с помощью специальной аппаратуры регистрируют ее продвижение по пищеварительному тракту. Радиоизотопные методы нашли также широкое применение в оценке желчевыделения, состояния печени, поджелудочной и слюнных желез.
Исследование процессов гидролиза и всасывания питательных веществ. Конечный результат дает много сведений о процессах гидролиза и всасывания питательных веществ из пищеварительного тракта. О нарушении гидролиза можно судить по наличию в кале непереваренных компонентов пищи.
Гидролиз и всасывание углеводов можно исследовать следующим образом: испытуемый натощак выпивает определенное количество раствора крахмала. Отсутствие увеличения содержания глюкозы в крови (по сравнению с эффектом перорального введения того же количества глюкозы) указывает на нарушение гидролиза полисахаридов. Содержание глюкозы в крови, учитываемое в течение нескольких часов после приема внутрь дисахаридов (мальтозы, сахарозы, лактозы и трегалозы), дает возможность сделать выводы о недостаточности дисахаридазных систем поджелудочной железы и тонкой кишки. Пробы с пероральным или зондовым введением в кишку мономеров питательных веществ (жирные кислоты, моносахариды, аминокислоты) с радиоактивной меткой с последующим их определением в крови и кале позволяют количественно оценить процесс всасывания в тонкой кишке.
Таким образом, современная физиология располагает методическими приемами, позволяющими исследовать пищеварительные функции на различных уровнях их организации, механизмы регуляции этих функций в норме и при патологии, и тем самым составляет основу функциональной диагностики клинической гастроэнтерологии.
9.3.5. Регуляция пищеварительных функций
9.3.5.1. Системные механизмы управления пищеварительной деятельностью. Рефлекторные механизмы
Натощак пищеварительный тракт находится в состоянии относительного покоя, для которого характерна периодическая функциональная активность (см. раздел 9.3.5.4). Прием пищи оказывает рефлекторное пусковое влияние на проксимальные отделы пищеварительного тракта: резко и кратковременно усиливаются секреция слюнных, желудочных и поджелудочной желез, желчевыделение, расслабляется желудок и снижается моторная активность проксимального отдела тонкой кишки.
Пусковые влияния стимулируют выделение секретов с высокой ферментативной активностью, обеспечиваемой накопленными в железах в период их относительного покоя ферментами (по И. П. Павлову — «запальный сок»).
|
|
|
Рис. 9.7. Механизмы регуляции секреции и моторики пищеварительного тракта. ПЖ — пищеварительные железы; ГМ — гладкие мышцы; ЭК — эндокринные клетки; Р — рецепторы; С и П — аксоны симпатических и парасимпатических преганглионарных нейронов; ПК — паракринные влияния; ЦНС — центральная нервная система. |
В дальнейшем секреция и моторика изменяют свой характер за счет корригирующих нервных, гуморальных и паракринных влияний по принципу обратной связи, формирующихся на основе рецепции содержимого, пищеварительного тракта его секреции и моторики. Следовательно, находящаяся в нем смесь пищевого содержимого с пищеварительными соками является объектом, в котором происходит пищеварительный процесс, одновременно параметры этой смеси -служат средством регуляции пищеварительного процесса путем изменения секреции, моторики и всасывания в пищеварительном тракте.
В полости рта осуществляется осознаваемая рецепция вкусов и запахов (благодаря проникновению пахучих веществ из полости рта к рецепторам носовых ходов), температуры, влажности, механических и некоторых других свойств пищи. В пищеварительном тракте осуществляется подсознательная рецепция. Здесь контролируются объем и консистенция принятой пищи, степень наполнения органа и давление в нем, наличие недостаточно размельченных кусочков пищи, рН, осмотическое давление, температура, концентрация питательных веществ и продуктов их гидролиза, активность некоторых пищеварительных ферментов и концентрация некоторых их фрагментов.
Рецепция осуществляется нервными рецепторными окончаниями, заложенными в стенке пищеварительного тракта (механо-, хемо-, осмо-, терморецепторы). Кроме того, афферентными сигналами служат всосавшиеся в кровь продукты гидролиза питатель ных веществ (глюкоза, некоторые аминокислоты), а также регуляторные пептиды, высвобождаемые различными клетками-продуцентами, находящимися в стенке желудка и кишечника. В этих клетках рецепторная функция совмещена с высвобождением регуляторных пептидов и аминов, которые затем с помощью различных механизмов влияют на клетки-мишени, реализующие пищеварительные функции.
Афферентные сигналы формируются и в органах-эффекторах в зависимости от их секреторной, моторной и всасывательной активности.
Афферентация в пищеварительном тракте по принципу отрицательной и положительной обратной связи имеет ряд специфических особенностей и важна в регуляции и саморегуляции всех пищеварительных функций, их системной, адаптивной по значению организации во времени и пространстве в зависимости от этапного и конечного результатов пищеварительного процесса.
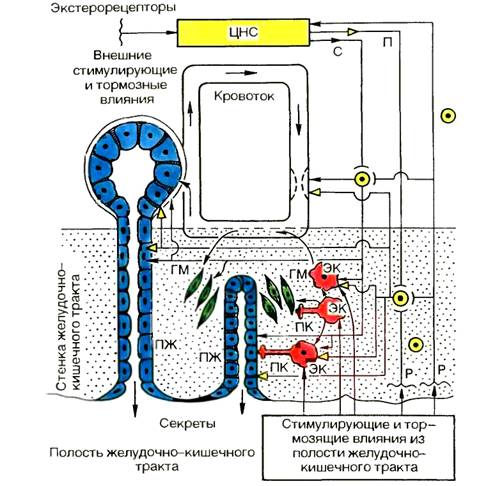
Механизмы эфферентных влияний на органы пищеварения сложны и различны. Еще И. П. Разенков (1948) считал, что пищеварительный тракт имеет три основных механизма регуляции секреторной и моторной функций (рис. 9.7), в разной мере представленных в его отделах: 1) центральные рефлекторные механизмы регулируют функции преимущественно начального отдела пищеварительного тракта, в дистальнее расположенных органах их роль снижается, при этом рефлекторная регуляция с экстеро- и интероцепторов осуществляется через рефлекторные дуги, замыкающиеся в ЦНС, экстра- и интрамуральных ганглиях; 2) гуморальные — наиболее выражены в «средней» части пищеварительного тракта — в регуляции деятельности желудка и двенадцатиперстной кишки, еще более — поджелудочной железы, желчеобразования и желчевыделения (гастродуоденопанкреатогепатобилиарный комплекс), где большую роль играют регуляторные пептиды, высвобождаемые эндокринными клетками самого пищеварительного тракта и приносимые кровотоком к гландулоцитам и миоцитам. Регуляторный пептид может поступать из продуцирующей его клетки в интерстициальную ткань, диффундировать в жидкости к расположенным рядом миоцитам, гландулоцитам и иннервирующим их нейронам; 3) локальные, т. е. местные, механизмы регуляции в наибольшей мере представлены в дистальных отделах — в деятельности тонкой и особенно толстой кишки.
Афферентная импульсная информация поступает от пищеварительного тракта в систему нейронов, расположенных в интра- и экстрамуральных ганглиях, спинном и головном мозге. Из них по эфферентным нервным путям импульсы следуют в органы пищеварительной системы. Нервная система пищеварительного тракта имеет симпатическую, парасимпатическую и метасимпатическую части. Их сплетения имеют рецепторные, вставочные и эфферентные нейроны. В окончаниях нейронов выделяются различные медиаторы, которые модулируют активность этих нейронов и клеток-эффекторов: гландулоцитов, миоцитов, энтероцитов. Перифе рические рефлекторные дуги обеспечивают не столько пусковые, сколько корригирующие и адаптационно-трофические влияния. Они важны в организации согласованных и сложных сокращений желудка, кишечника и сфинктеров.
Среди вегетативных нейронов, иннервирующих пищеварительный тракт, имеются не только холин- и адренергические, но и несколько типов пептидергических нейронов. В роли медиаторов в них выступают нейропептиды: например, симпатические преган-глионарные нейроны выделяют ацетилхолин (АХ), энкефалин и нейротензин; постганглионарные — норадреналин, АХ, вазоактивный интестинальный пептид (ВИП); парасимпатические преганглионарные нейроны — АХ и энкефалин, а постганглионарные — АХ, энкефалин и ВИП. В роли нейротрансмиттеров в желудке и кишечнике выступают также соматостатин, гастрин, нейротензин, субстанция Р, ВИП, холецистокинин (ХЦК). Медиаторами сенсорных нейронов также выступает ряд нейропептидов.
Каждому виду пищи соответствуют определенная моторика и секреция различных отделов пищеварительного тракта. Адаптация проявляется в соответствии объема, электролитного состава и спектра ферментов секретов пищеварительных желез принятой пище.
|
|
|
Рис 9.8.Фазы секреции главных пищеварительных желез. 1 — желудочная секреция при выраженной «мозговой фазе»; 2 — желудочная секреция при заторможенности «мозговой фазы»; 3 — секреция поджелудочной железы. |
Различают видовые и индивидуальные адаптации; последние делят на медленные, постепенно формируемые и фиксируемые на значительное время адаптации к длительным рационам питания, и быстрые (срочные), состоящие в приспособлении секреции ферментов и электролитов, моторики к определенному виду принятой пищи.
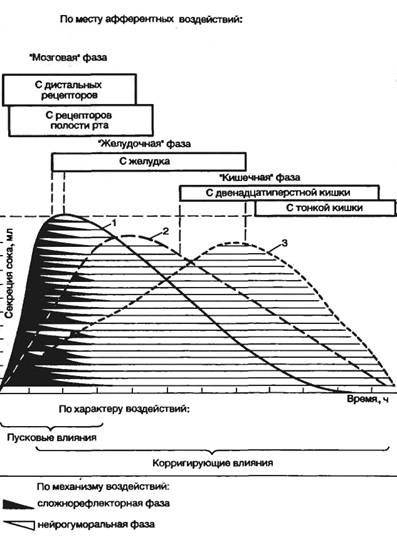
Фазы секреции пищеварительных желез. Образцом системного анализа механизмов регуляции пищеварительных функций явилось учение И. П. Павлова о фазах секреции главных пищеварительных желез, которое в настоящее время существенно дополнено и продолжает развиваться (рис. 9.8).
Секреция пищеварительных желез начинается с так называемой мозговой, или первой, фазы. Она осуществляется сложно-рефлекторно через ЦНС с участием условных и безусловных рефлексов. Секрецию стимулируют условно-рефлекторно вид, запах еще не принятой пищи и другие раздражители, связанные с ее приемом (обстановка, время, сервировка стола и др.); безусловно-рефлекторно — путем раздражения пищей рецепторов полости рта и пищевода. Вторая фаза секреции желез, возникающая при раздражении рецепторного аппарата желудка и высвобождении его гуморальных агентов, называется «желудочной». «Кишечной», или третьей, фазой называется секреция, опосредуемая интестинальными гормонами, раздражением рецепторов кишечника и всосавшимися из него питательными веществами.
Деление секреции на фазы (см. рис. 9.8) основано на нескольких принципах: по месту афферентации, механизму и характеру воздействия на пищеварительные железы. Учение о фазах секреции было сначала предложено для объяснения секреции желудочных
желез, но оно справедливо и для поджелудочной железы. Каждая из фаз имеет не только стимулирующий, но и тормозной компонент, что обеспечивает корригирующую роль динамического регуляторного контроля за секрецией. Учение о фазах секреции в принципе может быть перенесено и на системный анализ моторной деятельности пищеварительного тракта. Моторика тонкой кишки усиливается в раздражаемом и более дистальном ее сегменте, но тормозится в проксимальном ее участке. Это характерно и для моторики всего пищеварительного тракта и его секреции. Так, недостаточный гидролиз пищи в желудке задерживает эвакуацию из него пищевого содержимого. Если в силу тех или иных причин желудочная секреция в первую («мозговую») фазу заторможена (а она легко тормозится по многим причинам), то обычно удлиняется секреция в «желудочную» и «кишечную» фазы. В целом же недостаточность пищеварительного процесса в данном отделе пищеварительного тракта усиливает деятельность его последующих отделов, компенсируя запрограммированный уровень переработки пищи путем усиления секреции и увеличения времени пребывания пищевого содержимого в желудке и замедления транзита химуса по тонкой кишке. Эта схема регуляции имеет много вариантов в норме и часто нарушается при патологии.
9 .3.5.2. Роль регуляторных пептидов в деятельности пищеварительного тракта
В управлении пищеварительными функциями принимают участие пептиды и амины, которые продуцируются эндокринными клетками самого пищеварительного тракта. Эти клетки рассеяны в слизистой оболочке и пищеварительных железах и в совокупности составляют диффузную эндокринную систему. Продукты их деятельности называют гастроинтестинальными гормонами, энтеринами, регуляторными пептидами пищеварительного тракта. Это не только пептиды, но и амины. Некоторые из них продуцируются и нервными клетками. В первом случае эти биологически активные вещества выступают в роли гормонов (доставляются к органам-мишеням общим и региональным кровотоком) и парагормонов (диффундируют через интерстициальную ткань к рядом или недалеко расположенной клетке). Во втором случае эти вещества играют роль нейротрансмиттеров.
Открыто более 30 регуляторных пептидов пищеварительного тракта, некоторые из них существуют в нескольких изоформах, отличаясь числом аминогрупп и физиологической активностью. Выявлены клетки, продуцирующие данные пептиды и амины (табл. 9.1), а также клетки, в которых образуется не один, а несколько пептидов. Установлено, что один и тот же пептид может образовываться в различных клетках.
Гастроинтестинальные гормоны имеют широкий спектр физиологической активности, влияя на пищеварительные функции и вызывая общие эффекты. В пищеварительном тракте пептиды и амины стимулируют, тормозят, модулируют секрецию, моторику, всасывание, оказывают трофические влияния, в том числе воздействуя на пролиферативные процессы, например изменяют количество глан дулоцитов в слизистой оболочке желудка и поджелудочной железе, уменьшая или увеличивая их массу. Каждый из регуляторных пептидов вызывает несколько эффектов, один из которых часто является основным (табл. 9.2). Ряд пептидов выступает в роли рилизинг-факторов для других пептидов, которые вызывают изменения пищеварительных функций в таком регуляторном каскаде. Эффекты регуляторных пептидов зависят от их дозы, механизмов, с помощью которых была стимулирована функция. Сложны сочетанные влияния нескольких регуляторных пептидов, а также пептидов с влияниями автономной (вегетативной) нервной системы.
Регуляторные пептиды относятся к числу «короткоживущих» веществ (период полураспада несколько минут), вызываемые ими эффекты, как правило, значительно длительнее.
|
Таблица 9.1. Типы и локализация эндокринных клеток пищеварительного тракта и образуемые ими продукты |
|
Типы клеток |
Образуемые продукты |
Место расположения клетки | |||||
|
поджелудочная железа |
желудок |
тонкая кишка |
Толстая кишка | ||||
|
фун-дальная часть |
антраль-ная часть |
проксимальный отдел |
дис-тальный отдел | ||||
|
EC |
Серотонин, Вещество P,энкефалин |
Мало |
+ |
+ |
+ |
+ |
+ |
|
D |
Соматостатин |
+ |
+ |
+ |
+ |
Мало |
Мало |
|
B |
Инсулин |
+ |
- |
- |
- |
- |
- |
|
PP |
Панкреатический пептид(ПП) |
+ |
|||||
|
A |
Глюкагон |
+ |
- |
- |
- |
- |
- |
|
X |
Неизвестны |
- |
+ |
- |
- |
- |
- |
|
ECL |
Неизвестны(серотонин?гистамин?) |
- |
+ |
- |
- |
- |
- |
|
G |
Гастрин |
- |
- |
+ |
+ |
- |
- |
|
CCK |
Холецистокинин(ХЦК) |
- |
- |
- |
+ |
Мало |
- |
|
S |
Секретин |
- |
- |
- |
+ |
Мало |
- |
|
GIP |
Гастроингибирующий пептид(ГИП |
- |
- |
- |
+ |
Мало |
- |
|
M |
Мотилин |
- |
- |
- |
+ |
Мало |
- |
|
N |
Нейротизин |
- |
- |
- |
Мало |
+ |
Редко |
|
|
Иммунологически подобный глюкагону пептид, глицентин. |
- |
- |
- |
Мало |
+ |
+ |
|
GRP |
Гастрин-рилизинг-пептид |
- |
Мало |
+ |
+ |
- |
- |
|
VIP |
Вазоактивный интестиальный пептид (ВИП) |
||||||
|
Таблица 9.2. Основные эффекты влияния гастроинтестинальных гормонов на пищеварительные функции. |
|
Гормоны |
Эффекты (наиболее выраженные для человека) |
|
Гастрин |
Усиление секреции желудка (соляной кислоты и пепсиногена) и поджелудочной железы, гипертрофия слизистой оболочки желудка, усиление моторики желудка, тонкой и толстой кишки и желчного пузыря |
|
Секретин |
Увеличение секреции гидрокарбонатов поджелудочной железой, потенцирование действия холецистокинина (ХЦК) на поджелудочную железу, торможение секреции соляной кислоты в желудке и его моторики, усиление желчеобразования, секреции тонкой кишки |
|
Холецистокинин(ХЦК) |
Усиление моторики желчного пузыря и секреции ферментов поджелудочной железой, торможение секреции соляной кислоты в желудке и его моторики, усиление секреции в нем пепсиногена, моторики тонкой и толстой кишки, расслабление сфинктера печеночно-поджелудочной (ампулы Одни). Угнетение аппетита, гипертрофия поджелудочной железы |
|
Гастроингибирующий(желудочный, ингибирующий, пептид,ГИП или ЖИП) |
Глюкозозависимое усиление высвобождения поджелудочной железой инсулина, торможение секреции и моторики желудка путем снижения высвобождения гастрина, усиление кишечной секреции и торможение всасывания электролитов в тонкой кишке |
|
Мотилин |
Усиление моторики желудка и тонкой кишки, секреции пепсиногена желудком, секреции тонкой кишки |
|
Нейротензин |
Торможение секреции соляной кислоты желудком, усиление секреции поджелудочной железы, потенцирование эффектов секретина и ХЦК |
|
Панкреатический пептид(ПП) |
Антагонист ХЦК. Торможение секреции ферментов и гидрокарбонатов поджелудочной железой, усиление пролиферации слизистой оболочки тонкой кишки, поджелудочной железы и печени, релаксация желчного пузыря, усиление моторики желудка и тонкой кишки |
|
Энтероглюкагон |
Мобилизация углеводов, торможение секреции желудка и поджелудочной железы, моторики желудка и кишечника, пролиферация слизистой оболочки тонкой кишки (индукция гликогенолиза, липолиза, глюконеогенеза и кетогенеза) |
|
Пептид(УУ) |
Торможение секреции желудка, поджелудочной железы (различие эффектов в зависимости от дозы и объекта исследования) |
|
Вазоактивный интестинальный пептид(ВИП) |
Расслабление гладких мышц кровеносных сосудов, желчного пузыря, сфинктеров, торможение секреции желудка, усиление секреции гидрокарбонатов поджелудочной железой, кишечной секреции |
|
Гастрин-рилизинг-фактор |
Эффекты гастрина и усиление высвобождения ХЦК (и его эффекты) |
|
Химоденин Субстанция Р |
Стимуляция секреции поджелудочной железой химотрипсиногена |
|
Энкефалин |
Торможение секреции ферментов поджелудочной железой и желудком |
Концентрация пептидов в крови натощак колеблется в небольших пределах, прием пищи вызывает повышение концентрации ряда пептидов на разное время. Относительное постоянство содержания пептидов крови обеспечивается балансом поступления пептидов в кровоток с их ферментативной деградацией, небольшое количество их выводится из крови в составе секретов и экскретов, связывается белками крови. Деградация полипептидов ведет к образованию более простых олигопептидов, которые обладают большей или меньшей, иногда качественно измененной активностью. Дальнейший гидролиз пептидов приводит к потере их активности. В основном деградация пептидов происходит в почках и печени. Регуляторные пептиды пищеварительного тракта совместно с центральными и периферическими механизмами обеспечивают адаптивный характер и интеграцию пищеварительных функций.
9.3.5.3. Кровоснабжение и функциональная активность пищеварительного тракта
Напряженная деятельность органов пищеварения возможна лишь при интенсивном их кровоснабжении. При относительном функциональном покое пищеварительного тракта натощак в него поступает 15—20 % общего кровотока. После приема пищи кровоток в пищеварительном тракте может увеличиваться в несколько раз.
Кровоток в желудке и тонкой кишке примерно одинаков (30 мл/мин/100 г ткани), в толстой кишке он ниже. В стенке пищеварительного тракта преобладающая часть кровотока в соответствии с энергетическими затратами и функциональной активностью направляется в слизистую оболочку. После приема пищи ее кровоснабжение увеличивается в большей мере, чем в подслизистой основе и мышечном слое. Фильтрационная проницаемость капилляров пищеварительного тракта, особенно кишечника, примерно в 10 раз больше, чем в скелетных мышцах; после приема пищи в условиях вазодилатации фильтрационный коэффициент резко нарастает, что способствует транскапиллярному транспорту веществ.
Для пищеварительных желез характерен еще более высокий уровень кровотока. В состоянии относительного покоя кровоток в слюнных железах составляет 30—50 мл/мин/100 г ткани, в поджелудочной железе — 50—180 мл/мин/100 г, а при секреции и вазодилатации — 400 мл/мин/100 г. Соответственно в железах повышается и капиллярная проницаемость.
Реакция сердечно-сосудистой системы на прием пищи происходит в две фазы. В первую, длительностью 5—30 мин, рефлекторно повышаются минутный объем крови и артериальное кровяное давление, перераспределяется кровоток: уменьшается в скелетных мышцах и увеличивается в брыжеечной артерии. Вторая фаза в основном обусловлена местными и гуморальными механизмами, начинается через 30—90 мин после приема пищи и длится 3—7 ч. Во время второй фазы кровоток в скелетных мышцах снижен, а кровоснабжение органов пищеварения существенно увеличено.
Сопряжение уровня функциональной активности с величиной кровоснабжения органов пищеварения осуществляется с помощью нескольких механизмов: стимуляторы секреции и моторики являются и вазодилататорами (например, гистамин) или эти стимуляторы высвобождают из ткани сосудорасширяющие факторы (например, кинины); в результате стимуляции пищеварительных функций в органах образуются местно-действующие вазодилататоры, например калликреин; снижение Р02 и увеличение РС02,всасываемые пищевые вещества также обладают вазодилататорным действием (например, глюкоза в отношении сосудов кишечника и печени); освобождающиеся от секретов и уменьшающиеся в объеме железы снижают сопротивление в них кровотоку, и он нарастает.
В стенке пищеварительного тракта действует местный ауторегуляторный механизм обеспечения местного кровотока — способность сосудистого ложа поддерживать относительно постоянный кровоток, независимо от изменения системного кровяного давления, способность органа регулировать кровоснабжение в соответствии с функциональной активностью. Однако значительное снижение общего и регионального кровотока существенно уменьшает ее. Сокращения желудка и кишечника приводят к фазовым изменениям кровотока в них.
Регуляторные механизмы обеспечения местного кровотока в железах в основном те же, что и регуляции секреции желез. Это относится к большинству вегетативных нервов, регуляторных пептидов и аминов. Существенную роль в обеспечении местного кровотока желез играют вазодилататоры, образование которых в усиленно функционирующих железах повышается.
9.3.5.4. Периодическая деятельность органов пищеварения
В пищеварительном тракте условно выделяют несколько ритмов: базальные (секундные) ритмы с частотой 3—30 и более циклов в 1 мин; ритмы частотой 7—14 и 1 —14 и более циклов в сутки, приуроченные к темновому и световому периодам и связанные со временем приема пищи.
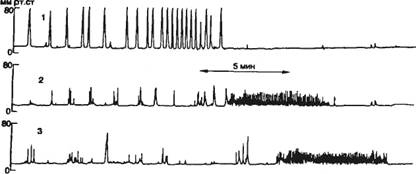
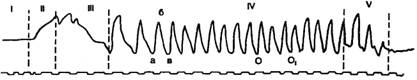
В лаборатории И. П. Павлова в хронических опытах на фистульных собаках В. Н. Болдыревым (1902) были открыты ритмы, названные периодической моторной деятельностью желудка. У человека через каждые 45—90 мин покоя — отсутствия сокращения желудка, наступает период его работы — сокращения, длящийся 20—50 мин. В моторной периодике принято различать 3 периода, или фазы: I — покоя; II — нерегулярных сокращений; III — регулярных сокращений (фронтальная активность).
В последние годы под термином «мигрирующий миоэлектрический (или моторный) комплекс» понимают перемещение мио-электрической, или сократительной, активности от желудка и
|
|
|
Рис. 9.9. Каудальное распространение моторной периодической активности пищеварительного тракта человека. Давление в антральной части желудка (1), двенадцатиперстной (2) и тощей (3) кишке во время фаз покоя и работы (с нерегулярными редкими и регулярными частыми сокращениями). |
двенадцатиперстной кишки до терминальной части подвздошной кишки (рис. 9.9).
Периодическая деятельность органов пищеварения проявляется не только в моторной активности пищеварительного тракта, но и секреции слюнных, желудочных, поджелудочных и кишечных желез, усилении желчеотделения и желчевыделения.
Синхронно с периодической деятельностью пищеварительного тракта изменяются интенсивность обмена веществ организма, температура тела, количество форменных элементов крови, концентрация гормонов и ферментативная активность крови, электроэнцефалографические показатели. Это свидетельствует о вовлечении в данный ритм периодической деятельности пищеварительного тракта всего организма.
Физиологическая роль периодической деятельности пищеварительного тракта полностью не установлена. Согласно одной из ранних гипотез, фазы работы периодической деятельности — сокращения желудка — вызывают чувство голода, поэтому ее назвали голодной периодикой. В классическом виде моторная периодика регистрируется только натощак. Прием пищи тормозит и существенно трансформирует ее, однако синхронность чувства голода и фазы работы периодики необязательна.
Допускают также, что физиологическим назначением периодической деятельности является транспорт из пищеварительных желез в кровь ферментов, изменяющих метаболизм организма. Еще одна гипотеза отводит периодической деятельности роль выведения из крови в полость пищеварительного тракта продуктов метаболизма. Достаточно популярна гипотеза о том, что периодическая деятельность своими перистальтическими волнами очищает пищеварительный тракт от остатков пищи и эндогенных веществ. Моторная периодика рассматривается как механизм предотвращения распространения кишечной микрофлоры в оральном направлении. Надо принять во внимание и то, что в ходе периодической секреции в пищеварительный тракт поступает значительное количество ферментов, которым затем предстоит осуществлять полостное и пристеночное пищеварение. Периодическая активность желез поддерживает их в состоянии некоторого оперативного покоя — готовности ответить выраженной секреторной реакцией на стимулы приема пищи.
В пищеварительный тракт в составе секретов и слущивающихся эпителиоцитов поступает много веществ (в том числе белков), представляющих большую пластическую и энергетическую ценность для организма. В пищеварительном тракте эти вещества подвергаются гидролизу, всасываются и утилизируются организмом. В связи с этим выдвинута гипотеза, согласно которой периодическая деятельность направлена на обеспечение эндогенного питания организма в условиях физиологического голода.
Перечень основных возможных назначений периодической деятельности пищеварительного тракта свидетельствует, что она осуществляет пищеварительные и непищеварительные функции в период между приемами пищи, но в несколько измененном виде. Она является одним из проявлений цикличности деятельности всего организма и участвует в обеспечении его гомеостаза. В основе периодической деятельности лежат периферические и центральные, рефлекторные и гормональные механизмы.
9.4. ПИЩЕВАРЕНИЕ В ПОЛОСТИ РТА И ГЛОТАНИЕ
Поступившая в рот пища раздражает рецепторы полости рта. Тактильные, температурные и болевые рецепторы расположены по всей слизистой оболочке рта, вкусовые — преимущественно во вкусовых почках сосочков языка. Они участвуют в формировании сладкого, кислого, горького и соленого вкуса. Различные зоны языка имеют различный набор рецепторов (см. главу 14).
Импульсы от вкусовых рецепторов по афферентным волокнам язычной ветви тройничного, лицевого и языкоглоточного нервов поступают в ЦНС. Эфферентные влияния возбуждают секрецию слюнных, желудочных и поджелудочной желез, желчевыделение, изменяют моторную деятельность пищевода, желудка, проксимального отдела тонкой кишки, влияют на кровоснабжение органов пищеварения, рефлекторно усиливают расходы энергии, необходимой для переработки и усвоения пищи (специфическое динамическое действие пищи; см. главу 10). Следовательно, несмотря на кратковременность пребывания пищи в полости рта ( в среднем 15—18 с), с ее рецепторов поступают пусковые влияния почти на весь пищеварительный тракт. Особенно важны раздражения рецепторов языка, слизистой оболочки рта и зубов в осуществлении пищеварительных процессов в самой полости рта. Здесь пища в процессе жевания измельчается, смачивается и перемешивается
|
|
|
Рис. 9.10. Кимограмма жевательного периода (по И. С. Рубинову). I — фаза покоя; II — фаза введения пищи в рот; III — ориентировочная фаза; IV — основная фаза; V — фаза формирования пищевого комка; а-b- опускание нижней челюсти, 6—в — подъем нижней челюсти; о—o1 — момент размалывания пищи. Внизу отметка времени 1 с. |
со слюной, растворяется (без чего невозможны оценка вкусовых качеств пищи и ее гидролиз); здесь же формируется ослизненный пищевой комок, предназначенный для глотания.
Пища принимается в виде кусков, смесей различного состава и консистенции или жидкостей. В зависимости от этого она либо подвергается механической и химической обработке в полости рта, либо сразу проглатывается. Процесс механической обработки пищи между верхними и нижними рядами зубов с помощью движения нижней челюсти относительно верхней называется жеванием. Жевательные движения осуществляются сокращениями жевательных и мимических мыщц, мыщц языка.
У взрослого человека имеется два ряда зубов. В каждом ряду с каждой стороны имеются резцы (2), клыки (1), малые (2) и большие коренные зубы (3). Резцы и клыки откусывают пищу, малые коренные зубы ее раздавливают, большие коренные зубы растирают. Резцы могут развивать давление на пищу 11—25 кг/см2, коренные зубы — 29—90 кг/см . Акт жевания осуществляется рефлекторно, имеет цепной характер, автоматизированные и произвольные компоненты.
Импульсы от рецепторов полости рта в основном по волокнам тройничного нерва передаются в сенсорные ядра продолговатого мозга, ядра зрительного бугра, оттуда — в кору большого мозга. От ствола мозга и зрительного бугра коллатерали отходят к ретикулярной формации. В регуляции жевания принимают участие двигательные ядра продолговатого мозга, красное ядро, черное вещество, подкорковые ядра и кора большого мозга. Совокупность управляющих жеванием нейронов различных отделов мозга называют центром жевания. Импульсы от него по двигательным волокнам тройничного нерва поступают к жевательным мышцам. Они осуществляют движения нижней челюсти вниз-вверх, вперед-назад и вбок. Мышцы языка, щек и губ перемещают пищевой комок в полости рта, подают и удерживают пищу между жевательными поверхностями зубов. В координации жевания большую роль играют импульсы от рецепторов жевательных мышц и зубов.
При регистрации жевания (мастикациография) выявляются следующие фазы (рис. 9.10): покоя, введения пищи в рот, ориентировочная, основная, формирования пищевого комка. Каждая из фаз и весь период жевания имеют различную длительность и характер, что зависит от свойств и количества пережевываемой пищи, возраста, аппетита, с которым принимается пища, индивидуальных особенностей, полноценности жевательного аппарата и механизмов его управления. Это определяет диагностическую ценность метода мастикациографии.
Слюна продуцируется тремя парами крупных слюнных желез и множеством мелких железок языка, слизистой оболочки неба и щек. Из желез по выводным протокам слюна поступает в полость рта. В зависимости от набора и интенсивности секреции разных гландулоцитов в железах они выделяют слюну разного состава. Околоушные и малые железы боковых поверхностей языка, содержащие большое количество серозных клеток, секретируют жидкую слюну с высокой концентрацией хлоридов натрия и калия и высокой активностью амилазы. Секрет поднижнечелюстной железы (смешанный) богат органическими веществами, в том числе муцином, содержит амилазу, но в меньшей концентрации, чем слюна околоушной железы. Слюна подъязычной железы (смешанная) еще более богата муцином, имеет выраженную щелочную реакцию, высокую фосфатазную активность. Секрет слизистых желез, расположенных в корне языка и неба, особенно вязок из-за высокой концентрации муцина. Здесь же есть и мелкие смешанные железы.
Из ацинусов желез секрет поступает в систему все укрупняющихся протоков, собирающихся в выводной проток, выносящий несколько измененную здесь (количество и состав) слюну в полость рта. Вне приема пищи у человека слюна выделяется для увлажнения полости рта в среднем со скоростью 0,24 мл/мин, при жевании — со скоростью 3—3,5 мл/мин в зависимости от вида пищи; при введении в рот лимонной кислоты (0,5 ммоль) — 7,4 мл/мин. За сутки выделяется 0,5—2,0 л слюны, около трети ее образуется околоушными железами.
Состав и свойства слюны. Смешанная слюна представляет собой вязкую, слегка опалесцирующую мутноватую жидкость с относительной плотностью 1,001—1,017, вязкостью 1,10—1,32 пуаза. Состав слюны зависит от скорости ее секреции и вида стимуляции саливации. Смешанная слюна имеет рН 5,8—7,4, рН слюны околоушных желез ниже (5,81), чем поднижнечелюстных (6,39). С увеличением скорости секреции рН слюны повышается до 7,8. Состав слюны сложен и меняется в зависимости от свойств принимаемой пищи, вида стимулятора слюновыделения (табл. 9.3). Муцин склеивает пищевые частицы в пищевой комок, который, будучи покрыт слизью, легче проглатывается. Этому способствует также пенообразование. Слизь слюны выполняет и защитную функцию, покрывая нежную слизистую оболочку рта и пищевода. Слюна содержит несколько ферментов: α-амилазу, α-глюкозидазу.
|
Таблица 9.3. Состав смешанной слюны человека |
|
Вещество |
Содержание, г/л |
Вещество |
Содержание, ммоль/л |
|
Вода |
994 |
Соли натрия |
6—23 |
|
Белки |
1,4—6,4 |
Соли калия |
14—41 |
|
Муцин |
0,8—6,0 |
Соли кальция |
1,2—2,7 |
|
Холестерин |
0,02—0,50 |
Соли магния |
0,1—0,5 |
|
Глюкоза |
0,1—0,3 |
Хлориды |
5—31 |
|
Аммоний |
0,01—0,12 |
Гидрокарбонаты |
2—13 |
|
Мочевая кислота |
0,005—0,030 |
Мочевина |
140—750 |
|
Фосфаты |
0,08—0,35 |
Гидролиз углеводов, осуществляемый с помощью этих ферментов, из-за кратковременности пребывания пищи в полости рта происходит в основном внутри пищевого комка уже в желудке. Действие карбогидраз слюны прекращается под влиянием кислой реакции желудочного сока. Активность протеолитических ферментов значительно ниже, а их роль в пищеварении взрослого человека невелика, однако эти ферменты имеют значение в санации полости рта. Так, мурамидаза (лизоцим) слюны обладает высокой бактерицидностью. Дезинфицирующее действие на содержимое полости рта оказывают протеиназы (саливаин, гландулаин и др.), РНКазы.
Значение слюны в пищеварении состоит в смачивании пищи, что способствует ее измельчению и гомогенизации при жевании; растворении питательных и вкусовых веществ, что важно для раздражения вкусовых рецепторов и действия ферментов слюны; ослизнения принятой и пережеванной пищи, что необходимо для формирования пищевого комка и облегченного его проглатывания.
Количество и состав слюны адаптированы к виду принимаемой пищи и режиму питания. На пищевые вещества выделяется более вязкая слюна, и ее тем больше, чем суше пища; на отвергаемые вещества и горечи — значительное количество жидкой слюны. Адаптация слюноотделения обеспечивается регуляторными воздействиями на слюнные железы.
Регуляция слюноотделения. Вне приема пищи небольшое количество слюны выделяют подъязычные, щечные и поднижнечелюстные железы человека. Прием пищи и связанные с ним факторы условно- и безусловнорефлекторно возбуждают слюноотделение. Латентный период слюноотделения зависит от силы пищевого раздражителя и возбудимости пищевого центра и составляет 1—30 с. Слюноотделение продолжается весь период еды и почти полностью прекращается вскоре после ее окончания. На стороне жевания слюны выделяется больше и с более высокой активностью амилазы, чем на противоположной стороне.
Возбуждение от рецепторов полости рта передается в ЦНС по афферентным волокнам тройничного, лицевого, языкоглоточного и блуждающего нервов. Импульсы достигают продолговатого мозга, других отделов мозга, включая кору большого мозга. Основной центр слюноотделения расположен в продолговатом мозге, сюда и в боковые рога верхних грудных сегментов спинного мозга поступают импульсы из вышерасположенных отделов мозга. К слюнным железам импульсы следуют по эфферентным парасимпатическим и симпатическим нервным волокнам.
Парасимпатическая иннервация поднижнечелюстной и подъязычной слюнных желез начинается от верхнего слюноотделительного ядра продолговатого мозга. Волокна преганглионарных нейронов в составе барабанной струны доходят до ганглиев этих желез, где переключаются на постганглионарные нейроны и по их аксонам достигают гландулоцитов. Преганглионарные волокна околоушных желез берут начало из нижнего слюноотделительного ядра продолговатого мозга, проходят в составе языкоглоточного нерва до ушного узла. Здесь расположены вторые нейроны, по аксонам которых в состав ушно-височного нерва импульсы достигают слюнных желез.
Под влиянием ацетилхолина, высвобождаемого окончаниями постганглионарных нейронов, выделяется большое количество жидкой слюны с высокой концентрацией электролитов и низкой концентрацией муцина.
Симпатическая иннервация слюнных желез осуществляется из боковых рогов II—IV грудных сегментов спинного мозга; отсюда волокна преганглионарных нейронов следуют в верхний шейный узел, где образуется контакт с постганглионарными нейронами. Их аксоны достигают слюнных желез.
Норадреналин, высвобождаемый окончаниями постганглионарных нейронов, вызывает выделение небольшого количества густой слюны, усиливает образование в железах ферментов и муцина. Одновременное раздражение парасимпатических нервов усиливает секреторный эффект. У человека симпатические нервы в наибольшей мере усиливают секрецию поднижнечелюстных слюнных желез.
Различия в секреции слюнных желез в ответ на прием различной пищи объясняются изменениями частот импульсов по парасимпатическим и симпатическим нервным волокнам, которые могут быть одно- и разнонаправленными. Слюноотделение относится к числу легкотормозимых процессов. Секрецию слюны тормозят болевые раздражения, отрицательные эмоции, умственное напряжение и др.
Парасимпатическая денервация слюнных желез вызывает их гиперсекрецию, это так называемая паралитическая секреция (максимум ее обычно отмечается на 7—8-й день после операции).
Снижение секреции слюнных желез называется гипосаливацией (гипосиалия). Она может вызвать многие нарушения, способствовать развитию микрофлоры во рту и быть причиной скверного запаха изо рта (есть и другие причины этого явления). Длительное снижение слюноотделения может быть причиной трофических нарушений слизистой оболочки рта, десен, зубов. Избыточное слюноотделение — гиперсаливация (сиалорея, птиализм) — сопровождает многие патологические состояния.
Жевание завершается глотанием — переходом пищевого комка из полости рта в желудок. Глотание возникает в результате раздражения чувствительных нервных окончаний тройничного, гортанных и языкоглоточного нервов. По афферентным волокнам этих нервов импульсы поступают в продолговатый мозг, где расположен центр глотания. От него импульсы по эфферентным двигательным волокнам тройничного, языкоглоточного, подъязычного и блуждающего нервов достигают мышц, обеспечивающих глотание. Доказательством рефлекторного характера глотания служит то, что если обработать корень языка и глотку раствором кокаина и «выключить» таким образом их рецепторы, то глотание не осуществится. Деятельность бульбарного центра глотания координируется двигательными центрами среднего мозга, коры больших полушарий. Бульварный центр находится в тесной связи с центром дыхания, тормозя его при глотании, что предотвращает попадание пищи в воздухоносные пути.
Рефлекс глотания состоит из трех последовательных фаз: I—ротовой (произвольной); II—глоточной (быстрой, короткой непроизвольной); III — пищеводной (медленной, длительной непроизвольной).
Во время фазы I из пищевой пережеванной массы во рту формируется пищевой комок объемом 5—15 см; движениями языка он перемещается на его спинку. Произвольными сокращениями передней, а затем средней части языка пищевой комок прижимается к твердому небу и переводится на корень языка за передние дужки.
Во время фазы II раздражение рецепторов корня языка рефлекторно вызывает сокращение мышц, приподнимающих мягкое небо, что препятствует попаданию пищи в полость носа. Движениями языка пищевой комок проталкивается в глотку. Одновременно происходит сокращение мышц, смещающих подъязычную кость и вызывающих поднятие гортани, вследствие чего закрывается вход в дыхательные пути, что препятствует поступлению в них пищи. Переводу пищевого комка в глотку способствуют повышение давления в полости рта и снижение давления в глотке. Препятствуют обратному движению пищи в ротовую полость поднявшийся корень языка и плотно прилегающие к нему дужки. Вслед за поступлением пищевого комка в глотку происходит сокращение мыщц, суживающих ее просвет выше пищевого комка, вследствие чего он продвигается в пищевод. Этому способствует разность давления в полостях глотки и пищевода.
Перед глотанием глоточно-пищеводный сфинктер закрыт, во время глотания давление в глотке повышается до 45 мм рт. ст., сфинктер открывается, и пищевой комок поступает в начало пище вода, где давление не более 30 мм рт. ст. Первые две фазы акта глотания длятся около 1 с. Фазу II глотания нельзя выполнить произвольно, если в полости рта нет пищи, жидкости или слюны. Если механически раздражать корень языка, то произойдет глотание, которое произвольно остановить нельзя. В фазу II вход в гортань закрыт, что предотвращает обратное движение пищи и попадание ее в воздухоносные пути.
Фазу III глотания составляют прохождение пищи по пищеводу и перевод ее в желудок сокращениями пищевода. Движения пищевода вызываются рефлекторно при каждом глотательном акте. Продолжительность фазы III при глотании твердой пищи 8—9 с, жидкой 1—2 с. В момент глотания пищевод подтягивается к зеву и начальная его часть расширяется, принимая пищевой комок. Сокращения пищевода имеют волновой характер, возникают в верхней его части и распространяются в сторону желудка. Такой тип сокращений называется перистальтическим. При этом последовательно сокращаются кольцеобразно расположенные мышцы пищевода, передвигая перетяжкой пищевой комок. Перед ним движется волна пониженного тонуса пищевода (релаксационная). Скорость ее движения несколько больше, чем волны сокращения, и она достигает желудка за 1—2 с.
Первичная перистальтическая волна, вызываемая актом глотания, доходит до желудка. На уровне пересечения пищевода с дугой аорты возникает вторичная волна, вызываемая первичной волной. Вторичная волна также продвигает пищевой комок до кардиальной части желудка. Средняя скорость ее распространения по пищеводу 2—5 см/с, волна охватывает участок пищевода длиной 10— 30 см за 3—7 с. Параметры перистальтической волны зависят от свойств проглатываемой пищи. Вторичная перистальтическая волна может быть вызвана остатком пищевого комка в нижней трети пищевода, благодаря чему он переводится в желудок. Перистальтика пищевода обеспечивает глотание и вне содействия ему сил гравитации (например, при горизонтальном положении тела или вниз головой, а также в условиях невесомости у космонавтов).
Прием жидкости вызывает глотание, которое в свою очередь формирует релаксационную волну, и жидкость переводится из пищевода в желудок не за счет пропульсивного его сокращения, а с помощью гравитационных сил и повышения давления в полости рта. Лишь последний глоток жидкости завершается прохождением пропульсивной волны по пищеводу.
Регуляция моторики пищевода осуществляется в основном эфферентными волокнами блуждающего и симпатического нервов; большую роль играет его интрамуральная нервная система.
Вне глотания вход из пищевода в желудок закрыт нижним пищеводным сфинктером. Когда релаксационная волна достигает конечной части пищевода, сфинктер расслабляется и перистальтическая волна проводит через него пищевой комок в желудок. При наполнении желудка тонус кардии повышается, что предотвращает забрасывание содержимого желудка в пищевод. Парасимпатические волокна блуждающего нерва стимулируют перистальтику пищевода и расслабляют кардию, симпатические волокна тормозят моторику пищевода и повышают тонус кардии. Одностороннему движению пищи способствует острый угол впадения пищевода в желудок. Острота угла увеличивается при наполнении желудка. Клапанную роль выполняет губовидная складка слизистой оболочки в месте перехода пищевода в желудок, сокращения косых мышечных волокон желудка и диафрагмально-пищеводная связка.
При некоторых патологических состояниях тонус кардии снижается, перистальтика пищевода нарушается и содержимое желудка может забрасываться в пищевод. Это вызывает неприятное ощущение, называемое изжогой. Нарушением глотания является аэрофагия — избыточное заглатывание воздуха, что чрезмерно повышает внутрижелудочное давление, и человек испытывает дискомфорт. Воздух выталкивается из желудка и пищевода, часто с характерным звуком (отрыгивание).
Пищеварительными функциями желудка являются депонирование, механическая и химическая обработка пищи и постепенная порционная эвакуация содержимого желудка в кишечник. Пища, находясь в течение нескольких часов в желудке, набухает, разжижается, многие ее компоненты растворяются и подвергаются гидролизу ферментами слюны и желудочного сока.
Карбогидразы слюны действуют на углеводы пищи, находящиеся в центральной части пищевого содержимого желудка, куда еще не диффундировал желудочный сок, прекращающий действие карбогидраз. Ферменты желудочного сока действуют на белки пищевого содержимого в зоне непосредственного контакта со слизистой оболочкой желудка и на небольшом удалении от нее, куда диффундировал желудочный сок.
Глубина проникновения желудочного сока зависит от его количества и свойств, от характера принятой пищи. Вся масса пищи в желудке не смешивается с соком. По мере разжижения и химической обработки пищи ее слой, прилегающий к слизистой оболочке, движениями желудка перемещается в антральную часть, откуда пищевое содержимое эвакуируется в кишечник. Таким образом, пищеварение в полости желудка осуществляется некоторое время за счет слюны, но ведущее значение имеет секреторная и моторная деятельность самого желудка.
9.5.1. Секреторная функция желудка
Образование, состав и свойства желудочного сока. Желудочный сок продуцируется железами желудка, расположенными в его слизистой оболочке. Она покрыта слоем цилиндрического эпителия, клетки которого секретируют слизь и слабощелочную жидкость. Слизь секретируется в виде густого геля, который покрывает равномерным слоем всю слизистую оболочку.
На поверхности слизистой оболочки видны мелкие впадинки — желудочные ямки. Общее их количество достигает 3 млн. В каждую из них открываются просветы 3—7 трубчатых желудочных желез. Различают три вида желудочных желез: собственные железы желудка, кардиальные и пилорические.
Собственные железы желудка располагаются в области тела и дна желудка (фундальные). Фундальные железы состоят из трех основных типов клеток: главные клетки — секретирующие пепсиногены, обкладочные (париетальные, оксинтные гландулоциты) — соляную кислоту и добавочные — слизь. Соотношение разных типов клеток в железах слизистой оболочки различных отделов желудка неодинаково. Кардиальные железы расположены в кардиальном отделе желудка — это трубчатые железы, состоящие в основном из клеток, продуцирующих слизь. В пилорическом отделе железы практически не имеют обкладочных клеток. Пилорические железы выделяют небольшое количество секрета, нестимулируемое приемом пищи. Ведущее значение в желудочном пищеварении имеет желудочный сок, вырабатываемый фундальными железами.
За сутки желудок человека выделяет 2—2,5 л желудочного сока. Он представляет собой бесцветную прозрачную жидкость, содержащую соляную кислоту (0,3—0,5%) и поэтому имеющую кислую реакцию (рН 1,5—1,8). Величина рН содержимого желудка значительно выше, так как сок фундальных желез частично нейтрализуется принятой пищей.
В желудочном соке имеются многие неорганические вещества: вода (995 г/л), хлориды (5—6 г/л), сульфаты (10 мг/л), фосфаты (10—60 мг/л), гидрокарбонаты (0—1,2 г/л) натрия, калия, кальция, магния, аммиак (20—80 кг/л). Осмотическое давление желудочного сока выше, чем плазмы крови.
Обкладочные клетки продуцируют соляную кислоту одинаковой концентрации (160 ммоль/л), но кислотность выделяющегося сока варьирует за счет изменения числа функционирующих париетальных гландулоцитов и нейтрализации соляной кислоты щелочными компонентами желудочного сока. Чем быстрее секреция соляной кислоты, тем меньше она нейтрализуется и тем выше кислотность желудочного сока.
Синтез соляной кислоты в обкладочных клетках сопряжен с клеточным дыханием и является аэробным процессом; при гипоксии секреция кислоты прекращается. Согласно «карбоангидразной» гипотезе, ионы Н+ для синтеза соляной кислоты получаются в результате гидратации СО2 и диссоциации образовавшейся при этом Н2СО3. Этот процесс катализируется ферментом карбоангидразой. Согласно «редокс»-гипотезе, ионы Н+ для синтеза соляной кислоты поставляются митохондриальной дыхательной цепью, а транспорт ионов Н+ и С1- осуществляется за счет энергии окислительно-восстановительных цепей. «АТФазная» гипотеза утверждает, что для транспорта этих ионов используется энергия АТФ, а Н+ могут происходить из различных источников, в том числе поставляться карбоангидразой из фосфатной буферной системы.
Сложные процессы, завершающиеся синтезом и экструзией из обкладочных клеток соляной кислоты, включают в себя три звена: 1) реакции фосфорилирования—дефосфорилирования; 2) митохондриальную окислительную цепь, работающую в режиме помпы; т. е. переносящую протоны из матриксного пространства вовне; 3) Н+, К+-АТФазу секреторной мембраны, осуществляющую «перекачку» этих протонов из клетки в просвет желез за счет энергии АТФ.
Соляная кислота желудочного сока вызывает денатурацию и набухание белков и тем самым способствует их последующему расщеплению пепсинами, активирует пепсиногены, создает кислую среду, необходимую для расщепления пищевых белков пепсинами; участвует в антибактериальном действии желудочного сока и регуляции деятельности пищеварительного тракта (в зависимости от рН его содержимого усиливается или тормозится нервными механизмами и гастроинтестинальными гормонами его деятельность).
Органические компоненты желудочного сока представлены азотсодержащими веществами (200—500 мг/л): мочевиной, мочевой и молочной кислотами, полипептидами. Содержание белка достигает 3 г/л, мукопротеидов — до 0,8 г/л, мукопротеаз — до 7 г/л. Органические вещества желудочного сока являются продуктами секреторной деятельности желудочных желез и обмена веществ в слизистой оболочке желудка, а также транспортируются через нее из крови. В числе белков особое значение для пищеварения имеют ферменты.
Главные клетки желудочных желез синтезируют несколько пепсиногенов, которые принято делить на две группы. Пепсиногены первой группы локализуются в фундальной части желудка, второй группы — в антральной части и начале двенадцатиперстной кишки. При активации пепсиногенов путем отщепления от них полипептида образуется несколько пепсинов. Собственно пепсинами принято называть ферменты класса протеаз, гидролизующие белки с максимальной скоростью при рН 1,5—2,0. Протеаза, названная гастриксином, имеет оптимальный для гидролиза белков рН 3,2— 3,5. Соотношение содержания пепсина и гастриксина в желудочном соке человека колеблется от 1:2 до 1:5. Эти ферменты различаются действием на разные виды белков.
Пепсины являются эндопептидазами, и основными продуктами их гидролитического действия на белки являются полипептиды (разрываются около 10% связей с освобождением аминокислот). Способность пепсинов гидролизовывать белки в широком диапазоне рН имеет большое значение для желудочного протеолиза, который происходит при разном рН в зависимости от объема и кислотности желудочного сока, буферных свойств и количества принятой пищи, диффузии кислого сока в глубь пищевого желудочного со держимого. Гидролиз белков происходит в непосредственной близости от слизистой оболочки. Проходящая перистальтическая волна «снимает» («слизывает») примукозальный слой, продвигает его к антральной части желудка, в результате чего к слизистой оболочке примыкает бывший более глубокий слой пищевого содержимого, на белки которого пепсины действовали при слабокислой реакции. Эти белки подвергаются гидролизу пепсинами в более кислой среде.
Важным компонентом желудочного сока являются мукоиды, продуцируемые мукоцитами поверхностного эпителия, шейки фундальных и пилорических желез (до 15 г/л). К мукоидам относится и гастромукопротеид (внутренний фактор Касла). Слой слизи толщиной 1—1,5 мм защищает слизистую оболочку желудка и называется слизистым защитным барьером желудка. Слизь — мукоидный секрет — представлена в основном двумя типами веществ — гликопротеинами и протеогликанами.
Сок, выделяемый разными участками слизистой оболочки желудка, содержит различное количество пепсиногена и соляной кислоты. Так, железы малой кривизны желудка продуцируют сок с более высокими кислотностью и содержанием пепсина, чем железы большой кривизны желудка.
Железы в пилорической части желудка выделяют небольшое количество сока слабощелочной реакции с большим содержанием слизи. Увеличение секреции происходит при местном механическом и химическом раздражении пилорической части желудка. Секрет пилорических желез обладает небольшой протеолитической, липолитической и амилолитической активностью. Существенного значения в желудочном пищеварении ферменты, обусловливающие эту активность, не имеют. Щелочной пилорический секрет частично нейтрализует кислое содержимое желудка, эвакуируемое в двенадцатиперстную кишку.
Показатели желудочной секреции имеют существенные индивидуальные, половые и возрастные различия. При патологии желудочная секреция может повышаться (гиперсекреция) или понижаться (гипосекреция), соответственно может меняться секреция соляной кислоты (гипер- и гипоацидность, отсутствие ее в соке — анацидность, ахлоргидрия). Меняется содержание пепсиногенов и соотношение их видов в желудочном соке.
Большое защитное значение имеет слизистый барьер желудка, разрушение которого может быть одной из причин повреждения слизистой оболочки желудка и даже глубже расположенных структур его стенки. Этот барьер повреждается при высокой концентрации в содержимом желудка соляной кислоты, алифатическими кислотами (уксусная, соляная, масляная, пропионовая) даже в небольшой концентрации, детергентами (желчные кислоты, салициловая и сульфосалициловая кислоты в кислой среде желудка), фосфолипазами, алкоголем. Длительный контакт этих веществ (при их относительно высокой концентрации) нарушает слизистый барьер и может привести к повреждению
|
|
|
Рис. 9.11. Кривые сокоотделения павловского желудочка на мясо, хлеб и молоко. |
слизистой оболочки желудка. Разрушению слизистого барьера и стимуляции секреции соляной кислоты способствует деятельность микроорганизмов Helicobacter pylori. В кислой среде и в условиях нарушенного слизистого барьера возможно переваривание элементов слизистой оболочки пепсином (пептический фактор язвообразования). Этому способствует также снижение секреции гидрокарбонатов и микроциркуляции крови в слизистой оболочке желудка.
Регуляция желудочной секреции. Вне пищеварения железы желудка выделяют небольшое количество желудочного сока. Прием пищи резко увеличивает его выделение. Это происходит за счет стимуляции желудочных желез нервными и гуморальными механизмами, составляющими единую систему регуляции. Стимулирующие и тормозные регуляторные факторы обеспечивают зависимость сокоотделения желудка от вида принимаемой пищи. Эта зависимость была впервые обнаружена в лаборатории И. П. Павлова в опытах на собаках с изолированным павловским желудочком, которым скармливалась различная пища. Объем и характер секреции во времени, кислотность и содержание в соке пепсинов определяются видом принятой пищи (рис. 9.11).
Стимуляция секреции соляной кислоты обкладочными клетками осуществляется непосредственно и опосредованно через другие механизмы. Непосредственно стимулируют секрецию соляной кислоты обкладочными клетками холинергические волокна блуждающих нервов, медиатор которых — ацетилхолин (АХ) — возбуждает М-холинорецепторы базолатеральных мембран гландулоцитов. Эффекты АХ и его аналогов блокируются атропином. Непрямая стимуляция клеток блуждающими нервами опосредуется также гастрином и гистамином.
Гастрин высвобождается из G-клеток, основное количество которых находится в слизистой оболочке пилорической части желудка. После хирургического удаления пилорической части желудочная секреция резко снижается. Высвобождение гастрина усиливается импульсами блуждающего нерва, а также местным механическим и химическим раздражением этой части желудка. Химическими стимуляторами G-клеток являются продукты переваривания белков — пептиды и некоторые аминокислоты, экстрактивные вещества мяса и овощей. Бели рН в антральной части желудка понижается, что обусловлено повышением секреции соляной кислоты железами желудка, то высвобождение гастрина уменьшается, а при рН 1,0 прекращается и объем секреции резко понижается. Таким образом, гастрин принимает участие в саморегуляции желудочной секреции в зависимости от величины рН содержимого антрального отдела. Гастрин в наибольшей мере стимулирует париетальные гландулоциты желудочных желез и увеличивает выделение соляной кислоты.
К стимуляторам обкладочных клеток желудочных желез относится и гистамин, образующийся в ECL-клетках слизистой оболочки желудка. Высвобождение гистамина обеспечивается гастрином. Гистамин стимулирует гландулоциты, влияя на Нг-рецепторы их мембран и вызывая выделение большого количества сока высокой кислотности, но бедного пепсином.
Стимулирующие эффекты гастрина и гистамина зависят от сохранности иннервации желудочных желез блуждающими нервами: после хирургической и фармакологической ваготомии секреторные эффекты этих гуморальных стимуляторов понижаются.
Желудочную секрецию возбуждают также всосавшиеся в кровь продукты переваривания белков.
Торможение секреции соляной кислоты вызывают секретин, ХЦК, глюкагон, ЖИП, ВИП, нейротензин, полипептид УУ, соматостатин, тиролиберин, энтерогастрон, АДГ, кальцитонин, окситоцин, простагландин ПГЕ2, бульбогастрон, кологастрон, серотонин (см. табл. 9.2). Высвобождение некоторых из них в соответствующих эндокринных клетках слизистой оболочки кишечника контролируется свойствами химуса. В частности, торможение желудочной секреции жирной пищей в большой мере обусловлено влиянием на железы желудка ХЦК. Повышение кислотности содержимого двенадцатиперстной кишки тормозит выделение соляной кислоты железами желудка. Торможение секреции осуществляется рефлекторно, а также вследствие образования гормонов двенадцатиперстной кишки.
Механизм стимуляции и торможения секреции соляной кислоты различными нейротрансмиттерами и гормонами неодинаков. Так, АХ усиливает секрецию кислоты обкладочными клетками путем активации мембранной Na+, К+-АТФазы, увеличения транспорта ионов Са2+ и эффектов повышенного внутриклеточного содержания цГМФ, высвобождения гастрина и потенцирования его влияния.
Гастрин усиливает секрецию соляной кислоты посредством гистамина, а также путем действия на мембранные рецепторы гастрина и усиления внутриклеточного транспорта ионов Са2+. Гистамин стимулирует секрецию обкладочных клеток через их мембранные Н2-рецепторы и систему аденилатциклаза (АЦ) — цАМФ.
Стимуляторами секреции пепсиногена главными клетками являются холинергические волокна блуждающих нервов, гастрин, гистамин, симпатические волокна, оканчивающиеся на β-адренорецепторах, секретин и ХЦК. Усиление секреции пепсиногенов главными клетками желудочных желез осуществляется несколькими механизмами. Среди них увеличение переноса ионов Са2+ в клетку и стимуляция Na+, К+-АТФазы; усиление внутриклеточного перемещения гранул зимогена, активация мембранной фосфорилазы, что усиливает их прохождение через апикальные мембраны, активация системы цГМФ и цАМФ.
Эти механизмы в неодинаковой мере активируются или тормозятся различными нейротрансмиттерами и гормонами, непосредственными и опосредованными влияниями их на главные клетки и секрецию пепсиногена. Показано, что гистамин и гастрин влияют на него опосредованно — усиливают секрецию соляной кислоты, а снижение рН содержимого желудка через местный холинергический рефлекс усиливает секрецию главных клеток. Описано и прямое стимулирующее влияние на них гастрина. В высоких дозах гистамин тормозит их секрецию. ХЦК, секретин и β-адреномиметики непосредственно стимулируют секрецию главных клеток, но тормозят секрецию обкладочных, что свидетельствует о существовании на них разных рецепторов регуляторных пептидов.
Стимуляция секреции слизи мукоцитами осуществляется холинергическими волокнами блуждающих нервов. Гастрин и гистамин умеренно стимулируют мукоциты, видимо, в связи с удалением слизи с их мембран при выраженной секреции кислого желудочного сока. Ряд ингибиторов секреции соляной кислоты — серотонин, соматостатин, адреналин, дофамин, энкефалин, простагландин ПГЕ2 — усиливает секрецию слизи. Полагают, что ПГЕ2 усиливает секрецию слизи названными веществами.
При приеме пищи и пищеварении в усиленно секретирующих железах желудка кровоток возрастает, что обеспечивается действием холинергических нервных механизмов, пептидов пищеварительного тракта и местных вазодилататоров. В слизистой оболочке кровоток нарастает интенсивнее, чем в подслизистой основе и мышечном слое желудочной стенки.
Фазы желудочной секреции. Нервные, гуморальные факторы и паракринные механизмы тонко регулируют секрецию желез желудка, обеспечивают выделение определенного количества сока, кислото- и ферментовыделение в зависимости от количества и качества принятой пищи, эффективности ее переваривания в желудке и тонкой кишке. Происходящую при этом секрецию принято делить на три фазы.
Начальная секреция желудка возникает рефлекторно в ответ на раздражение дистантных рецепторов, возбуждаемых видом и запахом пищи, всей обстановкой, связанной с ее приемом (услов-норефлекторные раздражения). Кроме того, секреция желудка возбуждается рефлекторно в ответ на раздражение принимаемой пищей рецепторов полости рта и глотки (безусловно-рефлекторные раздражения). Эти рефлексы обеспечивают пусковые влияния на железы желудка. Желудочную секрецию, обусловленную этими сложными рефлекторными влияниями, принято называть первой, или мозговой, фазой секреции (см. рис. 9.8).
Механизмы первой фазы секреции желудка были изучены в опытах на эзофаготомированных собаках с фистулой желудка. При кормлении такой собаки пища выпадает из пищевода и не поступает в желудок, однако через 5—10 мин после начала мнимого кормления начинает выделяться желудочный сок. Аналогичные данные были получены при исследовании людей, страдающих сужением пищевода и подвергшихся вследствие этого операции наложения фистулы желудка. Жевание пищи вызывало у людей выделение желудочного сока.
Рефлекторные влияния на желудочные железы передаются через блуждающие нервы. После их перерезки у эзофаготомированной собаки ни мнимое кормление, ни вид и запах пищи не вызывают секреции. Если раздражать периферические концы перерезанных блуждающих нервов, то отмечается выделение желудочного сока с высоким содержанием в нем соляной кислоты и пепсина.
В стимуляцию желудочных желез в первую фазу включен и гастриновый механизм. Доказательством этого служит увеличение содержания гастрина в крови людей при мнимом кормлении. После удаления пилорической части желудка, где продуцируется гастрин, секреция в первую фазу понижается.
Секреция в мозговую фазу зависит от возбудимости пищевого центра и может легко тормозиться при раздражении различных внешних и внутренних рецепторов. Так, плохая сервировка стола, неопрятность места приема пищи снижают и тормозят желудочную секрецию. Оптимальные условия приема пищи положительно влияют на желудочную секрецию. Прием в начале еды сильных пищевых раздражителей повышает желудочную секрецию в первую фазу.
На секрецию первой фазы наслаивается секреция второй фазы, которая называется желудочной, так как обусловлена влиянием пищевого содержимого в период его нахождения в желудке. Наличие этой фазы секреции доказывается тем, что вкладывание пищи в желудок через фистулу, вливание через нее или зонд растворов в желудок, раздражение его механорецепторов вызывают отделение желудочного сока. Объем секреции при этом в 2—3 раза меньше, чем при естественном приеме пищи. Это подчеркивает большое значение пусковых рефлекторных влияний, осуществляемых преимущественно в первую фазу на желудочные железы. Во вторую фазу железы желудка испытывают в основном корригирующие влияния. Эти влияния путем усиления и ослабления деятельности желез обеспечивают соответствие секреции количеству и свойствам пищевого желудочного содержимого, т. е. осуществляют коррекцию секреторной деятельности желудка. Сокоотделение при механическом раздражении желудка возбуждается рефлекторно с механорецепторов слизистой оболочки и мышечного слоя стенки желудка. Секреция резко уменьшается после перерезки блуждающих нервов. Кроме того, механическое раздражение желудка, особенно его пилорической части, приводит к высвобождению из G-клеток гастрина.
Повышение кислотности содержимого антральной части желудка тормозит высвобождение гастрина и снижает желудочную секрецию. В фундальной части желудка кислотность его содержимого рефлекторно усиливает секрецию, особенно выделение пепсиногена. Определенное значение в реализации желудочной фазы секреции имеет гистамин, значительное количество которого образуется в слизистой оболочке желудка.
Мясной бульон, капустный сок, продукты гидролиза белков при введении в тонкую кишку вызывают выделение желудочного сока. Нервные влияния с рецепторов кишечника на железы желудка обеспечивают секрецию в третью, кишечную, фазу. Возбуждающие и тормозные влияния из двенадцатиперстной и тощей кишки на железы желудка осуществляются с помощью нервных и гуморальных механизмов, корригирующих секрецию. Нервные влияния передаются с механо- и хеморецепторов кишечника. Стимуляция желудочных желез в кишечную фазу является прежде всего результатом поступления в двенадцатиперстную кишку недостаточно физически и химически обработанного содержимого желудка. В стимуляции желудочной секреции принимают участие всосавшиеся в кровь продукты гидролиза питательных веществ, особенно белков. Эти вещества могут возбуждать железы желудка опосредованно через гастрин и гистамин, а также непосредственно действуя на желудочные железы.
Торможение желудочной секреции в ее кишечную фазу вызывается рядом веществ в составе кишечного содержимого, которые по убывающей силе тормозного действия расположены в следующем порядке: продукты гидролиза жира, полипептиды, аминокислоты, продукты гидролиза крахмала, Н+ (рН ниже 3 оказывает сильное тормозное действие).
Высвобождение в двенадцатиперстной кишке секретина и ХЦК под влиянием поступившего в кишечник содержимого желудка и образовавшихся продуктов гидролиза питательных веществ тормозит секрецию соляной кислоты, но усиливает секрецию пепсиногена. Желудочную секрецию тормозят и другие кишечные гормоны из группы гастронов и глюкагон, а также серотонин.
Влияние пищевых режимов на желудочную секрецию. В экспериментах на животных И. П. Павловым с сотрудниками, а затем И. П. Разенковым с сотрудниками показано, что секреция желудочных желез значительно изменяется в зависимости от характера питания. При длительном (30-40 дней) употреблении пищи, содержащей большое количество углеводов (хлеб, овощи), секреция уменьшается (в основном во вторую и третью фазы). Если животное длительный срок (30-60 дней) принимает пищу, богатую белками, например мясо, то секреция увеличивается, в особенности во вторую и третью фазы. При этом меняются не только объем и динамика во времени желудочной секреции, но и ферментативные свойства желудочного сока. А. М. Уголевым экспериментально установлено, что длительный прием растительной пищи повышает активность желудочного сока по отношению к белкам растительного происхождения («фитолитическая активность»), а преобладание в пищевом рационе животных белков повышает способность желудочного сока гидролизовать их («зоолитическая активность»). Это связано с изменением кислотности сока и соотношения в нем видов и свойств пепсинов.
9.5.2. Моторная функция желудка
Во время и в первые минуты после приема пищи желудок расслабляется — наступает пищевая рецептивная релаксация желудка (рис. 9.12), которая способствует депонированию пищи в желудке и его секреции. Спустя некоторое время в зависимости от вида пищи сокращения усиливаются, при этом наименьшая сила сокращения отмечается в кардиальной части желудка и наибольшая — в антральной. Сокращения желудка начинаются на большой кривизне в непосредственной близости от пищевода, где находится кардиальный водитель ритма. Второй водитель ритма локализован в пилорической части желудка.
При баллонной гастрографии (рис. 9.13) регистрируется три типа волн сокращений желудка: I — однофазные волны низкой амплитуды, давление колеблется от 1—2 до 5—10 мм рт. ст., длительностью 5—20 с; II — однофазные волны большой амплитуды, давление составляет 40—80 мм рт. ст., длительностью 12— 60 с; III — сложные волны, возникают на фоне меняющегося исходного давления. Волны I и II типов носят перистальтический характер, поддерживают определенный тонус желудка, обеспечивают смешивание пищи с желудочным соком в непосредственной близости к слизистой оболочке желудка. Средняя частота этих волн 3 в 1 мин. В центральной части желудка содержимое не перемешивается, поэтому пища, принятая в разное время, располагается в желудке слоями (стратификация). Волны III типа характерны для пилорической части желудка, носят пропульсивный характер и участвуют в эвакуации содержимого в двенадцатиперстную кишку.
При регистрации внутрижелудочного давления методом открытых катетеров выявляются сокращения желудка двух типов: фазовые (тип А) и тонические (тип Б). Волны типа А быстрые, перистальтические, продолжительностью 10—20 с с частотой около 3 в 1 мин, волны типа Б — медленные, тонические, длительностью до 2 мин. Волны типа А бывают двух видов, первые имеют амплитуду и колебание давления от 1 до 15 мм рт. ст., у вторых амплитуда и колебание давления 16—30 мм рт. ст. Тонические волны (тип Б) могут сочетаться и не сочетаться с фазовыми.
|
|
|
Рис. 9.12. Влияние глотания (указано стрелкой) на моторику пищевода (1), кардиального сфинктера (2), фундального (3) и антрального (4) отделов желудка собаки. Высота вертикальных линий справа соответствует давлению 25 мм вод. ст. Нижняя кривая (5) — пневмограмма. |
|
|
|
Рис. 9.13. Три типа волн сокращений желудка здорового человека (I—III). а — сокращения желудка; б — пневмограмма. |
В наполненном пищей желудке возникают три основных вида движений: перистальтические волны, систолические сокращения пилорического отдела и тонические, уменьшающие размер полости дна и тела желудка. Частота перистальтических сокращений около 3 в 1 мин; они распространяются от кардиальной части желудка к пилорической со скоростью около 1 см/с, быстрее по большой, чем по малой кривизне, длятся около 11/2 с. В пилорической части скорость распространения перистальтической волны увеличивается до 3—4 см/с.
После приема пищи и в зависимости от ее вида параметры моторной деятельности желудка приобретают характерную динамику. В течение первого часа перистальтические волны слабые, в дальнейшем они усиливаются (в пилорическом отделе увеличиваются их амплитуда и скорость распространения), проталкивая пищу к выходу из желудка. Давление в пилорическом отделе повышается до 10—25 см вод. ст., открывается сфинктер привратника (пилорический сфинктер), и порция желудочного содержимого переходит в двенадцатиперстную кишку. Оставшееся (большее) количество его возвращается в проксимальную часть пилорического отдела желудка. Такие движения желудка обеспечивают перемешивание и перетирание (фрикционный эффект) пищевого содержимого, его гомогенизацию. Характер, интенсивность, временная динамика моторики зависят от количества и вида пищи, от эффективности ее переваривания в желудке и кишечнике, обеспечивается регуляторными механизмами.
Регуляция моторики желудка. Раздражение блуждающих нервов и выделение АХ усиливают моторику желудка: увеличивают ритм и силу сокращений, ускоряют движение перистальтических волн. Влияния блуждающих нервов могут оказывать и тормозной эффект: рецептивная релаксация желудка, снижение тонуса пилорического сфинктера. Раздражение симпатических нервов и активация α-адренорецепторов тормозят моторику желудка: уменьшают ритм и силу его сокращений, скорость движения перистальтической волны. Описаны и стимулирующие α- и β-адренорецепторные влияния (например, на пилорический сфинктер). Двунаправленные влияния осуществляются пептидергическими нейронами. Названные типы влияний осуществляются рефлекторно при раздражении рецепторов рта, пищевода, желудка, тонкой и толстой кишки. Замыкание рефлекторных дуг осуществляется на различных уровнях ЦНС, в периферических симпатических ганглиях и интрамуральной нервной системе.
В регуляции моторики желудка велико значение гастроинтестинальных гормонов. Моторику желудка усиливают гастрин, мотилин, серотонин, инсулин, а тормозят — секретин, ХЦК, глюкагон, ЖИП, ВИП. Механизм их влияний на моторику прямой (непосредственно на мышечные пучки и миоциты) и опосредованный через интрамуральные нейроны. Моторика желудка зависит от уровня его кровоснабжения и сама влияет на него, изменяя сопротивление кровотоку при сокращениях желудка.
9.5.3. Эвакуация содержимого желудка в двенадцатиперстную кишку
Скорость эвакуации пищи из желудка зависит от многих факторов: объема, состава и консистенции (степени измельченности, разжиженности), величины осмотического давления, температуры и рН содержимого желудка, градиента давления между полостями пилорического отдела желудка и двенадцатиперстной кишки, состояния сфинктера привратника, аппетита, с которым принималась пища, состояния водно-солевого гомеостаза и ряда других причин. Пища, богатая углеводами, при прочих равных условиях быстрее эвакуируется из желудка, чем богатая белками. Жирная пища эвакуируется из него с наименьшей скоростью. Жидкости начинают переходить в кишку сразу после их поступления в желудок.
Время полной эвакуации смешанной пищи из желудка здорового взрослого человека составляет 6—10 ч.
Эвакуация из желудка растворов и пережеванной пищи происходит по экспоненте, а эвакуация жиров экспоненциальной зависимости не подчиняется. Скорость и дифференцированность эвакуации определяются согласованной моторикой гастродуоденального комплекса, а не только деятельностью сфинктера привратника, выполняющего в основном роль клапана.
Скорость эвакуации пищевого содержимого желудка имеет широкие индивидуальные различия, принимаемые за норму. Дифференцированность эвакуации в зависимости от вида принятой пищи выступает как закономерность без существенных индивидуальных особенностей и нарушается при различных заболеваниях органов пищеварения.
Регуляция скорости эвакуации содержимого желудка. Осуществляется рефлекторно при активации рецепторов желудка и двенадцатиперстной кишки. Раздражение механорецепторов желудка ускоряет эвакуацию его содержимого, а двенадцатиперстной кишки — замедляет. Из химических агентов, действующих на слизистую оболочку двенадцатиперстной кишки, значительно замедляют эвакуацию кислые (рН меньше 5,5) и гипертонические растворы, 10 % раствор этанола, глюкоза и продукты гидролиза жира. Скорость эвакуации зависит также от эффективности гидролиза питательных веществ в желудке и тонкой кишке; недостаточность гидролиза замедляет эвакуацию. Следовательно, желудочная эвакуация «обслуживает» гидролитический процесс в двенадцатиперстной и тонкой кишке и в зависимости от хода его с различной скоростью «загружает» основной «химический реактор» пищеварительного тракта — тонкую кишку.
Регуляторные влияния на моторную функцию гастродуоденального комплекса передаются с интеро- и экстероцепторов через ЦНС и короткие рефлекторные дуги, замыкающиеся в экстра- и интрамуральных ганглиях. В регуляции эвакуаторного процесса принимают участие гастроинтестинальные гормоны, влияющие на моторику желудка и кишечника, изменяющие секрецию главных пищеварительных желез и через нее — параметры эвакуируемого желудочного содержимого и кишечного химуса.
Рвотой называется непроизвольный выброс содержимого пищеварительного тракта через рот (иногда и нос). Рвоте часто предшествует неприятное ощущение тошноты. Рвота начинается сокращениями тонкой кишки, в результате чего часть ее содержимого антиперистальтическими волнами выталкивается в желудок. Через 10—20 с происходят сокращения желудка, раскрывается кардиальный сфинктер, после глубокого вдоха сильно сокращаются мышцы брюшной стенки и диафрагмы, вследствие чего содержимое в момент выдоха выбрасывается через пищевод в полость рта; рот широко раскрывается, и из него удаляются рвотные массы. Их попадание в воздухоносные пути обычно предотвращено остановкой дыхания, изменением положения надгортанника, гортани и мягкого неба.
Рвота имеет защитное значение и возникает рефлекторно в результате раздражения корня языка, глотки, слизистой оболочки желудка, желчных путей, брюшины, коронарных сосудов, вестибулярного аппарата (при укачивании), мозга. Рвота может быть обусловлена действием обонятельных, зрительных и вкусовых раздражителей, вызывающих чувство отвращения (условнорефлекторная рвота). Ее также вызывают некоторые вещества, действующие гуморально на нервный центр рвоты. Эти вещества могут быть эндогенными и экзогенными.
Центр рвоты расположен на дне IV желудочка в ретикулярной формации продолговатого мозга. Он связан с центрами других отделов мозга и центрами других рефлексов. Импульсы к центру рвоты поступают от многих рефлексогенных зон. Эфферентные импульсы, обеспечивающие рвоту, следуют к кишечнику, желудку и пищеводу в составе блуждающего и чревного нервов, а также нервов, иннервирующих брюшные и диафрагмальные мышцы, мышцы туловища и конечностей, что обеспечивает основные и вспомогательные движения (в том числе и характерную позу). Рвота сопровождается изменением дыхания, кашлем, потоотделением, слюноотделением и другими реакциями.
9.6. ПИЩЕВАРЕНИЕ В ТОНКОЙ КИШКЕ
В обеспечении начального этапа пищеварения большая роль принадлежит процессам, происходящим в двенадцатиперстной кишке. Натощак ее содержимое имеет слабощелочную реакцию (рН 7,2—8,0). При переходе в кишку порций кислого содержимого желудка реакция содержимого двенадцатиперстной кишки становится кислой, но затем она сдвигается к нейтральной за счет поступающих в кишку щелочных секретов поджелудочной железы,
|
|
|
Рис. 9.14. Динамика величины рН содержимого антральной части желудка и двенадцатиперстной кишки. |
тонкой кишки и желчи (рис. 9.14), которые прекращают действие желудочного пепсина. В инактивации пепсина велика роль желчи. У человека рН содержимого двенадцатиперстной кишки колеблется в пределах 4—8,5. Чем выше его кислотность, тем больше выделяется сока поджелудочной железы, желчи и кишечного секрета, замедляется эвакуация содержимого желудка в двенадцатиперстную кишку и ее содержимого в тощую кишку. По мере продвижения по двенадцатиперстной кишке пищевое содержимое смешивается с поступающими в кишку секретами, ферменты которых уже в двенадцатиперстной кишке осуществляют гидролиз питательных веществ. Особенно велика в этом роль сока поджелудочной железы.
9.6.1. Секреция поджелудочной железы
Образование, состав и свойства сока поджелудочной железы. Основную массу поджелудочной железы (80—85 %) составляют экзокринные элементы, среди которых 80—95 % приходится на ацинозные (ацинарные) клетки; эти клетки секретируют ферменты (и небольшое количество неферментных белков); центроацинозные и протоковые клетки секретируют воду, электролиты, слизь; из протоков компоненты смешанного секрета частично реабсорбируются.
Поджелудочная железа человека натощак выделяет небольшое количество секрета. При поступлении пищевого содержимого из желудка в двенадцатиперстную кишку поджелудочная железа человека выделяет сок со средней скоростью 4,7 мл/мин. За сутки выделяется 1,5—2,5 л сока сложного состава (табл. 9.4).
|
Таблица 9.4. Состав сока поджелудочной железы |
|
Вещество |
Содержание,г/л |
Вещество |
Содержание,ммоль/л |
|
Вода |
987 |
Гидрокарбонаты |
25-150 |
|
Общий азот |
0,8-1,0 |
Хлориды |
4-129 |
|
Общий белок |
1,9-3,4 |
Соли калия |
6-9 |
|
Глюкоза |
0,08-0,18 |
Соли натрия |
139-143 |
|
Ферменты1 |
Соли кальция |
1,1-2,5 | |
1-К ферментам поджелудочной железы относятся α-амилаза, прокарбоксипептидазы (А1,А2,B1,B2), трипсиногены (1, 2, 3), химотрипсиноген, проэластаэы (1, 2), колипазы (1, 2), профосфолипаза А2, нуклеазы.
Сок представляет собой бесцветную прозрачную жидкость со средним содержанием воды 987 г/л. Щелочная среда сока (рН 7,5—8,8) обусловлена наличием в нем гидрокарбонатов (до 150 ммоль/л). Концентрация гидрокарбонатов в соке изменяется прямо пропорционально скорости секреции. В соке содержатся хлориды натрия и калия; между концентрацией гидрокарбонатов и хлоридов обратная зависимость. Гидрокарбонаты сока поджелудочной железы участвуют в нейтрализации и ощелачивании кислого пищевого содержимого желудка в двенадцатиперстной кишке. В соке отмечается значительная концентрация белка, основную часть которого составляют ферменты.
Сок поджелудочной железы богат ферментами, которые синтезируются в ацинозных панкреоцитах. Ферменты поджелудочного сока переваривают все виды питательных веществ. Амилаза, липаза и нуклеаза секретируются поджелудочной железой в активном состоянии, а протеазы — в виде зимогенов.
Трипсиноген сока поджелудочной железы в двенадцатиперстной кишке под действием ее фермента энтерокиназы превращается в трипсин. Последующую активацию трипсиногена вызывает трипсин. Активация состоит в отщеплении от трипсиногена гексапептида под действием энтерокиназы и трипсина при рН 6,8—8,0. Процесс ускоряется в присутствии ионов Са2+.
Химотрипсиноген активируется трипсином. Трипсин и химотрипсин (а также панкреатопептидаза, или эластаза) расщепляют преимущественно внутренние пептидные связи белков. Эти ферменты действуют и на высокомолекулярные полипептиды, в результате чего образуются низкомолекулярные пептиды и аминокислоты. В составе сока поджелудочной железы выделяется некоторое количество ингибитора трипсина.
Поджелудочная железа синтезирует прокарбоксипептидазы А и В, проэластазы и профосфолипазу. Они активируются трипсином с образованием соответствующих ферментов: карбоксипептидаз А и В, эластаз и фосфолипазы. Сок поджелудочной железы богат α-амилазой, расщепляющей полисахариды до ди- и моносахаридов. На производные нуклеиновых кислот действуют рибо- и дезоксирибонуклеазы. Панкреатическая липаза расщепляет жиры, в основном триглицериды, до моноглицеридов и жирных кислот. На липиды действуют также фосфолипаза А2 и эстераза.
Поджелудочная железа секретирует профермент — панкреатическую фосфолипазу, которая активируется трипсином. Поскольку триглицериды нерастворимы в воде, липаза действует только на поверхности жира. Чем больше суммарная площадь поверхности контакта жира и липазы, тем активнее идет его гидролиз. Поэтому эмульгирование жира имеет огромное значение для его переваривания. Эмульгирование обеспечивается желчью, точнее — ее желчными кислотами и их солями. Размеры частиц жира 0,2—5,0 мкм. Активность липазы повышает также фермент колипаза. Она связывается с липазой в присутствии желчных солей и снижает оптимум рН действия фермента с 9 до 6—7, а также способствует адсорбции липазы на слизистой оболочке кишки.
Повышают активность липазы присутствие и ионов Са2+. Под действием липаз осуществляется обычно неполный гидролиз три-глицеридов; при этом образуется смесь из моноглицеридов (около 50 %), жирных кислот и глицерина (40 %), ди- и триглицеридов (3—10%).
Регуляция секреции поджелудочной железы. Секреция поджелудочной железы регулируется нервными и гуморальными механизмами.
Нервная регуляция. И. П. Павлов показал, что раздражение блуждающего нерва вызывает выделение большого количества сока поджелудочной железы, богатого ферментами. Холинергические волокна блуждающих нервов с помощью АХ действуют на М-холинорецепторы панкреацитов. Затем высвобождающиеся ион Са2+ и комплекс ГЦ — цГМФ в роли вторичных мессенджеров стимулируют секрецию панкреоцитами ферментов и гидрокарбонатов. Холинергические нейроны, кроме того, потенцируют секреторные эффекты секретина и ХЦК. Хирургическая ваготомия существенно снижает секрецию поджелудочной железы.
Симпатические волокна, иннервирующие поджелудочную железу через β-адренорецепторы, тормозят ее секрецию, усиливают синтез органических веществ в ней. Адренергические эффекты снижения секреции обеспечиваются также уменьшением кровоснабжения поджелудочной железы путем сужения кровеносных сосудов через их α-адренорецепторы.
Торможение секреции вызывают болевые раздражения, сон, напряженная физическая и умственная работа и др.
Поджелудочная железа имеет также пептидергическую иннервацию. Окончания этих нейронов выделяют ряд нейропептидов, одни из которых стимулируют, другие — тормозят секрецию поджелудочной железы.
Гуморальная регуляция. Первым открытым (и на званным гормоном) явился секретин — стимулятор обильного сокоотделения и секреции гидрокарбонатов. Высвобождение этого гормона в кровь S-клетками двенадцатиперстной кишки происходит при действии на ее слизистую оболочку перешедшего в кишку кислого желудочного содержимого. Секретин стимулирует секрецию в большей мере через соответствующие мембранные рецепторы и вторичные мессенджеры АЦ — цАМФ центроацинозные и протоковые клетки, в меньшей мере — ацинозные клетки, поэтому выделяется секрет с высокой концентрацией гидрокарбонатов и низкой ферментативной активностью.
Вторым гормоном, усиливающим секрецию поджелудочной железы, является холецистокинин (ХЦК). Высвобождение гормона в кровь из ССК-клеток слизистой оболочки двенадцатиперстной и тощей кишки происходит под влиянием пищевого химуса (особенно продуктов начального гидролиза пищевых белков и жиров, углеводов, некоторых аминокислот). Стимулируют высвобождение ХЦК присутствие ионов Са2+ и снижение рН в двенадцатиперстной кишке.
ХЦК действует преимущественно на ацинусы поджелудочной железы, поэтому выделяющийся в ответ на стимуляцию этим гормоном сок богат ферментами. Вторичными мессенджерами являются ионы Са2+ и комплекс ГЦ — цГМФ. Одновременное действие на железу секретина и ХЦК (при приеме пищи) усиливает их стимуляторный эффект. Секретин и ХЦК применяются в клинике как стимуляторы секреции при диагностике заболеваний поджелудочной железы. Пептид химоденин стимулирует секрецию химотрипсиногена.
Секреция поджелудочной железы усиливается также гастрином, серотонином, инсулином, бомбезином, солями желчных кислот. Тормозят выделение поджелудочного сока глюкагон, соматостатин, вазопрессин, вещество Р, АКТГ, энкефалин, кальцитонин, ЖИП, ПП, УУ. ВИП может возбуждать и тормозить секрецию поджелудочной железы.
Фазы секреции поджелудочной железы. Секреция сока поджелудочной железы резко усиливается через 2—3 мин после приема пищи и продолжается 6—14 ч. От количества и качества пищи зависят объем, состав выделяющегося сока, динамика выделения. Чем выше кислотность пищевого содержимого желудка, поступающего в двенадцатиперстную кишку, тем больше выделяется сока поджелудочной железы и тем больше гидрокарбонатов в его составе. Поэтому кривая секреции поджелудочной железы в некоторой мере повторяет кривую желудочного сокоотделения. Отличия кривых секреции желудка и поджелудочной железы определяются в основном буферными свойствами пищи, которая частично нейтрализует кислоту желудочного сока, и скоростью эвакуации содержимого желудка в двенадцатиперстную кишку.
Фазы секреции поджелудочной железы при стимуляции ее приемом пищи те же, что были описаны для желудочной секреции, но в отличие от них более выражены гормональные влияния на
|
|
|
Рис. 9.15. Секреция поджелудочного сока у собаки при еде мяса, хлеба и молока. |
поджелудочную железу, особенно в кишечную фазу. Секреция имеет характерную динамику, зависящую от вида принятой пищи (рис. 9.15).
Первая, или мозговая, фаза секреции поджелудочной железы обусловлена видом, запахом пищи и другими раздражителями, связанными с приемом пищи (условнорефлекторные раздражения), а также воздействиями на рецепторы слизистой оболочки рта, жеванием и глотанием (безусловно-рефлекторные раздражения). Нервные импульсы, возникающие в рецепторах, достигают продолговатого мозга и затем по волокнам блуждающего нерва поступают к железе и вызывают ее секрецию.
У человека с фистулой протока поджелудочной железы наблюдали условнорефлекторное выделение сока поджелудочной железы через 2—3 мин после того, как испытуемому говорили о пище, которую ему дадут. В реализации первой фазы секреции принимают участие и регуляторные пептиды, высвобождение которых стимулировалось рефлекторными механизмами блуждающих нервов.
Вторая, или желудочная, фаза характеризуется тем, что секреция во время нее стимулируется и поддерживается путем ваговагального рефлекса с механо- и хеморецепторов желудка и с помощью гастрина.
С переходом желудочного содержимого в двенадцатиперстную кишку начинается третья, или кишечная, фаза секреции поджелудочной железы. В эту фазу секреция стимулируется ваговагальным дуоденопанкреатическим рефлексом, но ведущее значение имеет высвобождение в кровь секретина и ХЦК. Высвобождение их происходит при действии на слизистую оболочку двенадцатиперстной кишки кислого ее содержимого. Чем больше свободных ионов Н+, тем больше высвобождается секретина и тем выше объем сока поджелудочной железы и секреция гидрокарбонатов. Гидрокарбонаты связывают ионы Н+, что приводит к повышению рН среды и уменьшает высвобождение секретина и соответственно объем сока и секрецию гидрокарбонатов. Секреция ферментов в кишечную фазу стимулируется ХЦК и рефлекторно за счет ваговагального рефлекса.
В кишечную фазу велика роль саморегуляции секреции поджелудочной железы по принципу отрицательной обратной связи в зависимости от свойств содержимого двенадцатиперстной кишки. Установлено, что отведение сока поджелудочной железы из двенадцатиперстной кишки вызывает гиперсекрецию поджелудочной железы, обратное введение сока в кишку тормозит эту секрецию. Введение в кишку гидрокарбонатов снижает объем секреции, концентрацию и дебит (выделение) гидрокарбонатов в составе сока. Введение сока поджелудочной железы в двенадцатиперстную кишку особенно выраженно тормозит секрецию поджелудочной железой ферментов. При этом повышение триптической активности химуса двенадцатиперстной кишки тормозит секрецию протеаз, повышение амилолитической активности химуса тормозит секрецию амилазы, повышенная липолитическая активность в наибольшей мере тормозит секрецию панкреатической липазы. Тормозные эффекты ферментов снимаются или снижаются их специфическими ингибиторами и пищевыми субстратами (Г. Ф. Коротько).
Таким образом, свойства секрета поджелудочной железы в кишечную фазу в большой мере определяются соотношением в химусе двенадцатиперстной кишки ферментов и гидролизуемых ими субстратов: избыток ферментов селективно тормозит их секрецию, избыток субстрата снимает эти тормозные влияния, и продукты гидролиза субстрата стимулируют секрецию соответствующих ферментов поджелудочной железой. Этот механизм направлен на срочную адаптацию секреции ферментов поджелудочной железы к виду принятой пищи. Его реализация обеспечивается М-холинергическими и (β-адренергическими влияниями, ХЦК, секретином.
В целом нервные влияния при приеме пищи обеспечивают пусковые воздействия на поджелудочную железу, в последующей коррекции ее секреции большую роль играют гуморальные механизмы. Однако высвобождение гормонов двенадцатиперстной кишкой и действие их на поджелудочную железу более выражены при сохраненной ее иннервации, что подчеркивает единство нервных и гуморальных механизмов регуляции секреции поджелудочной железы. При стимуляции секреции поджелудочной железы усиливается ее кровоснабжение, что важно для поддержания секреции на высоком уровне.
Секреция гуморально корригируется и всосавшимися питательными веществами. Эти влияния осуществляются непосредственно на панкреоциты, выражено влияние на них некоторых аминокислот и глюкозы, опосредуются через центральные нервные механизмы (например, гипоталамический и бульварный центры автономной нервной системы) и регуляторные пептиды.
Влияние пищевых режимов на секрецию поджелудочной железы. Прием пищи вызывает увеличение выделения всех ферментов в составе сока, но для разных видов пищи это увеличение выражено в разной мере. В случае приема пищи с большим содержанием углеводов в наибольшей мере увеличивается секреция амилазы, белков — трипсина и химотрипсина, прием жирной пищи вызывает секрецию сока с более высокой липолитической активностью. Спектр ферментов сока поджелудочной железы срочно адаптируется к виду принимаемой пищи во все три фазы секреции и особенно в кишечную фазу.
Существуют и медленные адаптации секреции ферментов в зависимости от длительного режима питания. Суть адаптации состоит в том, что поджелудочная железа синтезирует и секретирует больше того фермента, который гидролизует преобладающие в рационе питательные вещества. Преобладание в рационе белков повышает выделение в составе сока поджелудочной железы протеаз, преимущественное углеводное питание вызывает увеличение выделения с соком амилазы, а большое количество жира в рационе — выделение с соком липазы.
Эти изменения носят согласованный с изменениями функционирования других отделов пищеварительного тракта характер, составляя часть интегрированных адаптации всего пищеварительного конвейера.
9.6.2. Желчеотделение и желчевыделение
Желчь, ее участие в пищеварении. Желчь образуется в печени, и ее участие в пищеварении многообразно. Желчь эмульгирует жиры, увеличивая поверхность, на которой осуществляется их гидролиз липазой; растворяет продукты гидролиза липидов, способствует их всасыванию и ресинтезу триглицеридов в энтероцитах; повышает активность ферментов поджелудочной железы и кишечных ферментов, особенно липазы. При выключении желчи из пищеварения нарушается процесс переваривания и всасывания жиров и других веществ липидной природы. Желчь усиливает гидролиз и всасывание белков и углеводов.
Желчь выполняет и регуляторную роль, являясь стимулятором желчеобразования, желчевыделения, моторной и секреторной деятельности тонкой кишки, пролиферации и слущивания эпителиоцитов (энтероцитов). Желчь способна прекращать действие желудочного сока, не только снижая кислотность желудочного содержимого, поступившего в двенадцатиперстную кишку, но и путем инактивации пепсина. Желчь обладает бактериостатическими свойствами. Немаловажной является ее роль во всасывании из кишечника жирорастворимых витаминов, холестерина, аминокислот и солей кальция.
У человека за сутки образуется 1000—1800 мл желчи (около 15 мл на 1 кг массы тела). Процесс образования желчи — желчеотделение (холерез) — осуществляется непрерывно, а поступление желчи в двенадцатиперстную кишку — желчевыделение (холекинез) — периодически, в основном в связи с приемом пищи. Натощак в кишечник желчь почти не поступает, она направляется в желчный пузырь, где при депонировании концентрируется и несколько изменяет свой состав, поэтому принято говорить о двух видах желчи — печеночной и пузырной (табл. 9.5).
Состав и образование желчи. Желчь является не только секретом, но и экскретом. В ее составе выводятся различные эндогенные и экзогенные вещества. Это определяет сложность состава желчи. В желчи содержатся белки, аминокислоты, витамины и другие вещества. Желчь обладает небольшой ферментативной активностью; рН печеночной желчи 7,3—8,0. При прохождении по желчевыводящим путям и нахождении в желчном пузыре жидкая и прозрачная золотисто-желтого цвета печеночная желчь (относительная плотность 1,008—1,015) концентрируется (всасываются вода и минеральные соли), к ней добавляется муцин желчных путей и пузыря, и желчь становится темной, тягучей, увеличивается ее относительная плотность (1,026—1,048) и снижается рН (6,0—7,0) за счет образования солей желчных кислот и всасывания гидрокарбонатов.
Основное количество желчных кислот и их солей содержится в желчи в виде соединений с гликоколом и таурином. Желчь человека содержит гликохолевых кислот около 80 % и таурохолевых — около 20 %. Прием пищи, богатой углеводами, увеличивает содержание гликохолевых кислот, в случае преобладания в диете белков увеличивается содержание таурохолевых кислот. Желчные кислоты и их соли определяют основные свойства желчи как пищеварительного секрета.
Желчные пигменты являются экскретируемыми печенью продуктами распада гемоглобина и других производных порфиринов. Основным желчным пигментом человека является билирубин — пигмент красно-желтого цвета, придающий печеночной желчи характерную окраску. Другой пигмент — биливердин (зеленого цвета) — в желчи человека содержится в следовых количествах, а появление его в кишечнике обусловлено окислением билирубина.
В желчи содержится комплексное липопротеиновое соединение, в состав которого входят фосфолипиды, желчные кислоты, холестерин, белок и билирубин. Это соединение играет важную роль в транспорте липидов в кишечник и принимает участие в печеночно-кишечном кругообороте и общем метаболизме организма.
Желчь состоит из трех фракций. Две из них образуются ге-патоцитами, третья — эпителиальными клетками желчных протоков. От общего объема желчи у человека на первые две фракции приходится 75 %, на долю третьей — 25%. Образование первой фракции связано, а второй — не связано напрямую с образованием желчных кислот. Образование третьей фракции желчи опреде
|
Таблица 9.5. Состав желчи |
|
Компоненты |
Печеночная желчь |
Пузырная желчь |
|
Вода, г/л |
950—980 |
850—920 |
|
Сухой остаток, г/л: |
||
|
соли жирных кислот |
10,0—11,0 |
30—100 |
|
жирные кислоты |
1,0 |
3,0—12,0 |
|
билирубин |
2,0 |
5,0—20,0 |
|
холестерин |
0,5—1,0 |
3,0—9,0 |
|
лецитин |
0,4 |
1,0—4,0 |
|
неорганические соли |
8,0— 8,5 |
6,0—6,5 |
|
Ионы, ммоль/л: |
||
|
Na+ |
145 |
130 |
|
К+ |
5 |
12 |
|
Са2+ |
2,5 |
10 |
|
С1- |
100 |
25 |
|
НСО3 |
28 |
10 |
ляется способностью эпителиальных клеток протоков секретировать жидкость с достаточно высоким содержанием гидрокарбонатов и хлора, осуществлять реабсорбцию воды и электролитов из канальцевой желчи.
Основной компонент желчи — желчные кислоты — синтезируются в гепатоцитах. Из тонкой кишки всасывается в кровь около 85—90 % желчных кислот, выделившихся в кишку в составе желчи. Всосавшиеся желчные кислоты с кровью по воротной вене транспортируются в печень и включаются в состав желчи. Остальные 10—15 % желчных кислот выводятся из организма в основном в составе кала. Эта потеря желчных кислот восполняется их синтезом в гепатоцитах.
В целом образование желчи происходит путем активного и пассивного транспорта веществ из крови через клетки и межклеточные контакты (вода, глюкоза, креатинин, электролиты, витамины, гормоны и др.), активной секреции компонентов желчи (желчные кислоты) гепатоцитами и обратного всасывания воды и ряда веществ из желчных капилляров, протоков и желчного пузыря (рис. 9.16). Ведущая роль в образовании желчи принадлежит секреции.
Регуляция желчеобразования. Желчеобразование осуществляется непрерывно, но интенсивность его изменяется за счет регуляторных влияний. Усиливают желчеобразование акт еды, принятая пища. Рефлекторно изменяется желчеобразование при раздражении интероцепторов пищеварительного тракта, других внутренних органов и условнорефлекторном воздействии.
Парасимпатические холинергические нервные волокна (воздействия) усиливают, а симпатические адренергические — снижают желчеобразование. Имеются экспериментальные данные об усилении желчеобразования под влиянием симпатической стимуляции.
|
|
|
Рис. 9.16. Желчеобразование и его регуляция. |
К числу гуморальных стимуляторов желчеобразования (холеретиков) относится сама желчь. Чем больше желчных кислот поступает из тонкой кишки в кровоток воротной вены (портальный кровоток) тем больше их выделяется в составе желчи, но меньше желчных кислот синтезируется гепатоцитами. Если поступление в портальный кровоток желчных кислот уменьшается, то дефицит их восполняется усилением синтеза желчных кислот в печени. Секретин усиливает секрецию желчи, выделение в ее составе воды и электролитов (гидрокарбонатов). Слабее стимулируют желчеобразование глюкагон, гастрин, ХЦК, простагландины.
Действие различных стимуляторов желчеобразования различно. Например, под влиянием секретина увеличивается в основном объем желчи, под влиянием блуждающих нервов, желчных кислот повышаются ее объем и выделение органических компонентов, высокое содержание в пище полноценных белков увеличивает выделение и концентрацию этих веществ в составе желчи. Желчеобразование усиливают многие продукты животного и растительного происхождения. Соматостатин уменьшает желчеобразование.
Желчевыделение. Движение желчи в желчевыделительном аппарате обусловлено разностью давления в его частях и в двенадцатиперстной кишке, состоянием сфинктеров внепеченочных желчных путей. В них выделяют следующие сфинктеры: в месте слияния пузырного и общего печеночного протока (сфинктер Мирисси), в шейке желчного пузыря (сфинктер Люткенса) и концевом отделе общего желчного протока и сфинктер ампулы, или Одди. Тонус мышц этих сфинктеров определяет направление движения желчи. Давление в желчевыделительном аппарате создается секреторным давлением желчеобразования и сокращениями гладких мышц протоков и желчного пузыря. Эти сокращения согласованы с тонусом сфинктеров и регулируются нервными и гуморальными механизмами. Давление в общем желчном протоке колеблется от 4 до 300 мм вод. ст., а в желчном пузыре вне пищеварения составляет 60—185 мм вод. ст., во время пищеварения за счет сокращения пузыря поднимается до 200—300 мм вод. ст., обеспечивая выход желчи в двенадцатиперстную кишку через открывающийся сфинктер Одди.
Вид, запах пищи, подготовка к ее приему и собственно прием пищи вызывают сложное и неодинаковое у разных лиц изменение деятельности желчевыделительного аппарата, при этом желчный пузырь сначала расслабляется, а затем сокращается. Небольшое количество желчи через сфинктер Одди выходит в двенадцатиперстную кишку. Этот период первичной реакции желчевыделительного аппарата длится 7—10 мин. На смену ему приходит основной эвакуаторный период (или период опорожнения желчного пузыря), во время которого сокращение желчного пузыря чередуется с расслаблением и в двенадцатиперстную кишку через открытый сфинктер Одди переходит желчь, сначала из общего желчного протока, затем пузырная, а в последующем — печеночная.
Длительность латентного и эвакуаторного периодов, количество выделенной желчи зависят от вида принятой пищи. Сильными стимуляторами желчевыделения являются яичные желтки, молоко, мясо и жиры.
Рефлекторная стимуляция желчевыделительного аппарата и холекинеза осуществляется условно- и безусловно-рефлекторно при раздражении рецепторов рта, желудка и двенадцатиперстной кишки с участием блуждающих нервов.
Наиболее мощным стимулятором желчевыделения является ХЦК, вызывающий сильное сокращение желчного пузыря; гастрин, секретин, бомбезин (через эндогенный ХЦК) вызывают слабые сокращения, а глюкагон, кальцитонин, антихолецистокинин, ВИП, ПП тормозят сокращение желчного пузыря.
Кишечный сок представляет собой мутную, вязкую жидкость, является продуктом деятельности всей слизистой оболочки тонкой кишки, имеет сложный состав и разное происхождение. За сутки у человека выделяется до 2,5 л кишечного сока.
В криптах слизистой оболочки верхней части двенадцатиперстной кишки заложены дуоденальные, или бруннеровы, железы. Клетки этих желез содержат секреторные гранулы муцина и зимогена. По строению и функции бруннеровы железы похожи на пилорические. Сок бруннеровых желез представляет собой густую бесцветную жидкость слабощелочной реакции, обладающую не большой протеолитической, амилолитической и липолитической активностью. Кишечные крипты, или либеркюновы железы, заложены в слизистой оболочке двенадцатиперстной и всей тонкой кишки и окружают каждую ворсинку.
Секреторной способностью обладают многие эпителиоциты крипт тонкой кишки. Зрелые кишечные эпителиоциты развиваются из недифференцированных бескаемчатых энтероцитов, которые преобладают в криптах. Эти клетки обладают пролиферативной активностью и восполняют кишечные клетки, которые подвергаются десквамации с верхушек ворсинок. По мере движения к верхушке бескаемчатые энтероциты дифференцируются в абсорбирующие клетки ворсинок и бокаловидные клетки.
Кишечные эпителиоциты с исчерченной каемкой, или абсорбирующие клетки, покрывают ворсинку. Их апикальная поверхность образована микроворсинками с выростами клеточной оболочки, тонкими филаментами, формирующими гликокаликс, а также содержит многие кишечные ферменты, транслоцированные из клетки, где они были синтезированы. Ферментами богаты также расположенные в апикальной части клеток лизосомы.
Бокаловидные клетки называют одноклеточными железами. Переполненная слизью клетка имеет характерный вид бокала. Выделение слизи происходит через разрывы апикальной плазматической мембраны. Секрет обладает ферментативной, в том числе и протеолитической, активностью.
Энтероциты с ацидофильными гранулами, или клетки Панета, в зрелом состоянии также имеют морфологические признаки секреции. Их гранулы гетерогенны и выводятся в просвет крипт по типу мерокриновой и апокриновой секреции. Секрет содержит гидролитические ферменты. В криптах заложены также аргентаффинные клетки, выполняющие эндокринные функции.
Находящееся даже в изолированной от остального кишечника полости петли тонкой кишки содержимое представляет собой продукт многих процессов (в том числе десквамации энтероцитов) и двустороннего транспорта высоко- и низкомолекулярных веществ. Это, собственно, и есть кишечный сок.
Свойства и состав кишечного сока. При центрифугировании кишечный сок разделяется на жидкую и плотную части. Соотношение между ними изменяется в зависимости от силы и вида раздражений слизистой оболочки тонкой кишки.
Жидкая часть сока образована секретом, транспортируемыми из крови растворами неорганических и органических веществ и частично — содержимым разрушенных клеток кишечного эпителия. Жидкая часть сока содержит около 20 г/л сухого вещества. В числе неорганических веществ (около 10 г/л) хлориды, гидрокарбонаты и фосфаты натрия, калия, кальция. рН сока 7,2— 7,5, при усилении секреции достигает 8,6. Органические вещества жидкой части сока представлены слизью, белками, аминокислотами, мочевиной и другими продуктами обмена веществ.
Плотная часть сока — желтовато-серая масса, имеющая вид слизистых комков и включающая в себя неразрушенные эпителиальные клетки, их фрагменты и слизь — секрет бокаловидных клеток имеет более высокую ферментативную активность, чем жидкая часть сока (Г. К. Шлыгин).
В слизистой оболочке тонкой кишки происходит непрерывная смена слоя клеток поверхностного эпителия. Они образуются в криптах, затем продвигаются по ворсинкам и слущиваются с их верхушек (морфокинетическая, или морфонекротическая, секреция). Полное обновление этих клеток у человека совершается за 1—4—6 сут. Такой высокий темп образования и отторжения клеток обеспечивает достаточно большое их количество в кишечном соке (у человека за сутки отторгается около 250 г эпителиоцитов).
Слизь образует защитный слой, предотвращающий чрезмерное механическое и химическое воздействие химуса на слизистую оболочку кишки.В слизи высока активность пищеварительных ферментов.
Плотная часть сока обладает значительно большей ферментативной активностью, чем жидкая. Основная часть ферментов синтезируется в слизистой оболочке кишки, но некоторое их количество транспортируется из крови. В кишечном соке более 20 различных ферментов, принимающих участие в пищеварении.
Основная часть кишечных ферментов принимает участие в пристеночном пищеварении. Углеводы гидролизируются α-глюкозидазами, α-галактазидазой (лактаза), глюкоамилазой (γ-амилаза). К α-глюкозидазам относятся мальтаза и трегалаза. Мальтаза гидролизует мальтозу, а трегалаза — трегалозу на 2 молекулы глюкозы. α-Глюкозидазы представлены еще одной группой дисахаридаз, куда входят 2—3 фермента с изомальтазной активностью и инвертаза, или сахараза; с их участием образуются моносахариды.
Высокая субстратная специфичность кишечных дисахаридаз при их дефиците обусловливает непереносимость соответствующего дисахарида. Известны генетически закрепленные и приобретенные лактазная, трегалазная, сахаразная и комбинированные недостаточности. У значительной популяции людей, особенно народов Азии и Африки, выявлена лактазная недостаточность.
В тонкой кишке продолжается и завершается гидролиз пептидов. Аминопептидазы составляют основную часть пептидазной активности щеточной каймы энтероцитов и расщепляют пептидную связь между двумя определенными аминокислотами. Аминопептидазы завершают мембранный гидролиз пептидов, в результате чего образуются аминокислоты — основные всасывающиеся мономеры.
Кишечный сок обладает липолитической активностью. В пристеночном гидролизе липидов особое значение имеет кишечная моноглицеридлипаза. Она гидролизует моноглицериды с любой длиной углеводородной цепи, а также короткоцепочечные ди- и триглицериды, в меньшей мере — триглицериды со средней длиной цепи и эфиры холестерина.
Ряд пищевых продуктов содержит нуклеопротеиды. Их начальный гидролиз осуществляется протеазами, затем гидролизуются отщепленные от белковой части РНК и ДНК соответственно РНК и ДНКазами до олигонуклеотидов, которые при участии нуклеаз и эстераз деградируют до нуклеотидов. Последние атакуются щелочными фосфатазами и более специфичными нуклеотидазами с высвобождением всасываемых затем нуклеозидов. Фосфатазная активность кишечного сока очень высокая.
Ферментный спектр слизистой оболочки тонкой кишки и ее сока изменяется под влиянием определенных длительных режимов питания.
Регуляция кишечной секреции. Прием пищи, местное механическое и химическое раздражение кишки усиливают секрецию ее желез с помощью холинергических и пептидергических механизмов.
В регуляции кишечной секреции ведущее значение имеют местные механизмы. Механическое раздражение слизистой оболочки тонкой кишки вызывает увеличение выделения жидкой части сока. Химическими стимуляторами секреции тонкой кишки являются продукты переваривания белков, жиров, сок поджелудочной железы, соляная и другие кислоты. Местное воздействие продуктов переваривания питательных веществ вызывает отделение кишечного сока, богатого ферментами.
Акт еды существенно не влияет на кишечную секрецию, в то же время имеются данные о тормозных влияниях на нее раздражения антральной части желудка, модулирующих влияний ЦНС, о стимулирующем действии на секрецию холиномиметических веществ и тормозном влиянии холинолитических и симпатомиметических веществ. Стимулируют кишечную секрецию ГИП, ВИП, мотилин, тормозит соматостатин. Гормоны энтерокринин и дуокринин, вырабатываемые в слизистой оболочке тонкой кишки, стимулируют соответственно секрецию кишечных крипт (либеркюновы железы) и дуоденальных (бруннеровых) желез. В очищенном виде эти гормоны не выделены.
9.6.4. Полостное и пристеночное пищеварение в тонкой кишке
В тонкой кишке выражены полостное и пристеночное пищеварение, о которых сказано в разделе 9.2.2, но не исключено и внутриклеточное пищеварение.
Полостное пищеварение в тонкой кишке осуществляется за счет пищеварительных секретов и их ферментов, поступивших в полость тонкой кишки (секрет поджелудочной железы, желчь, кишечный сок).
Полостное пищеварение в тонкой кишке осуществляется как в жидкой фазе кишечного химуса, так и на границе фаз: на поверхности пищевых частиц, растительных волокон, сохраненных и разрушенных десквамированных энтероцитов, хлопьев (флокул), образовавшихся в результате взаимодействия кислого содержимого желудка и щелочного дуоденального химуса. В результате полостного пищеварения гидролизуются крупномолекулярные вещества и образуются в основном олигомеры. Последующий их гидролиз происходит в зоне, прилегающей к слизистой оболочке, и непосредственно на ней (рис. 9.17).
|
|
|
Рис. 9.17. Взаимоотношения внутриполостного и мембранного пищеварения (по А. М. Уголеву, 1982). А — схема последовательной деполимеризации пищевых субстратов в полости и на поверхности тонкой кишки; Б — фрагмент липопротеидной мембраны с адсорбированными и собственно кишечными ферментами; М — мембрана; MB — микроворсинки; АГ — апикальный гликокаликс; ЛГ — латеральный гликокаликс; C1, C2, С3 — субстраты; ПФ — панкреатические ферменты; ТСМ — транспортная система мембраны; РЦФ — регуляторные центры ферментов; КЦФ — каталитические центры ферментов: НФ — неферментные факторы. |
Из полости тонкой кишки вещества поступают в слой кишечной слизи. Этот слой, образованный в основном секретом бокаловидных клеток и фрагментами слущивающегося кишечного эпителия, удерживается на пласте энтероцитов их гликокаликсом и за счет высокой вязкости секрета. Слой кишечной слизи непрерывно наращивается со стороны эпителиоцитов и убывает со стороны полости кишки, т. е. все время обновляется. Он обладает более высокой ферментативной активностью, чем жидкое содержимое полости тонкой кишки.
В слое кишечной слизи адсорбированы ферменты из полости тонкой кишки (панкреатические и кишечные), из разрушенных энтероцитов и транспортированные в кишку из кровотока. Проходящие через слои слизи питательные вещества частично гидролизуются этими ферментами — «премембранное пищеварение» (Ю. М. Гальперин) и поступают в слой гликокаликса, где продолжается гидролиз питательных веществ по мере их транспорта в глубь пристеночного слоя. Продукты гидролиза поступают на апикальные мембраны энтероцитов, в которые встроены кишечные ферменты, осуществляющие собственно мембранное пищеварение (в основном гидролиз димеров до стадии мономеров). Следовательно, пристеночное пищеварение последовательно осуществляется в трех зонах: в слое слизи, гликокаликсе и на апикальных мембранах энтероцитов. Образовавшиеся в результате пищеварения мономеры всасываются в кровь и лимфу.
Полостное пищеварение в тонкой кишке имеет выраженный проксимодистальный градиент: гидролиз интенсивнее совершается в проксимальной, чем в дистальной, части тонкой кишки. Топография мембранного пищеварения несколько другая, но имеется проксимодистальный градиент распределения ферментов вдоль тонкой кишки.
Эпителиоциты в разных частях кишечной ворсинки морфологически и функционально неоднозначны. Так, от крипты к вершине ворсинки секреторная активность эпителиоцитов убывает. Верхняя часть ворсинок преимущественно реализует мембранный гидролиз дипептидов, участки, расположенные ближе к основанию ворсинок — гидролиз дисахаридов. О функциональной специализации кишечных эпителиоцитов свидетельствует и распределение свойств тонкой кишки от проксимального к дистальному ее отделам. Это касается и гидролиза различных питательных веществ, и активности кишечных ферментов, и всасывания различных компонентов химуса.
Проксимодистальные градиенты свойств тонкой кишки имеют существенные различия, зависят от возраста, питания, времени суток, приема пищи и т. д. Существует последовательность в гидролизе и всасывании продуктов гидролиза питательных веществ, сложные взаимные влияния гидролиза одних веществ на гидролиз других.
Регуляция полостного и пристеночного пищеварения. Регуляция полостного пищеварения осуществляется путем изменения секреции пищеварительных желез, продвижения химуса по тонкой кишке, интенсивности пристеночного пищеварения и всасывания.
Регуляция пристеночного пищеварения более сложна и во многом еще не изучена. Интенсивность пристеночного пищеварения зависит от полостного и, следовательно, от факторов, влияющих на него. На мембранное пищеварение влияют гормоны надпочечников (синтез и транслокация ферментов), диеты и другие факторы. Пристеночное пищеварение зависит также от моторики кишки, изменяющей переход веществ из химуса в исчерченную каемку, величины пор исчерченной каемки, ферментного состава в ней, сорбционных свойств мембраны (А. М. Уголев).
9.6.5. Моторная функция тонкой кишки
Моторика тонкой кишки обеспечивает перемешивание ее содержимого (химуса) с пищеварительными секретами, продвижение химуса по кишке, смену его слоя у слизистой оболочки, повышение внутрикишечного давления, способствующего фильтрации растворов из полости кишки в кровь и лимфу. Следовательно, моторика тонкой кишки способствует гидролизу и всасыванию питательных веществ.
Движение тонкой кишки происходит в результате координированных сокращений продольного и циркулярного слоев гладких мышц. Принято различать несколько типов сокращений тонкой кишки: ритмическая сегментация, маятникообразные, перистальтические (очень медленные, медленные, быстрые, стремительные), антиперистальтические и тонические. Первые два типа относятся к ритмическим, или сегментирующим, сокращениям.
Ритмическая сегментация обеспечивается преимущественно сокращениями циркулярного слоя мышечной оболочки. При этом содержимое кишки делится на части. Следующим сокращением образуется новый сегмент кишки, содержимое которого состоит из химуса двух половин бывших сегментов. Данными сокращениями достигаются перемешивание химуса и повышение давления в каждом сегменте.
Маятникообразные сокращения обеспечиваются продольными мышцами и участием в сокращении циркулярных мышц. При этом происходят перемещение химуса вперед — назад и слабое поступательное движение его в каудальном направлении. В верхних отделах тонкой кишки человека частота ритмических сокращений составляет 9-12, в нижних — 6-8 в минуту.
Перистальтическая волна, состоящая из перехвата и расширения тонкой кишки, продвигает химус в каудальном направлении. Одновременно вдоль кишки продвигается несколько перистальтических волн. Перистальтическая волна продвигается по кишке со скоростью 0,1—0,3 см/с, в проксимальных отделах она больше, чем в дистальных. Скорость стремительной (пропульсивной) волны 7—21 см/с.
При антиперистальтических сокращениях волна движется в обратном (оральном) направлении. В норме тонкая кишка, как и желудок, антиперистальтически не сокращаются (это характерно для рвоты).
Тонические сокращения могут иметь локальный характер или перемещаться с очень малой скоростью. Тонические сокращения суживают просвет кишки на большом ее протяжении.
Исходное (базальное) давление в полости тонкой кишки составляет 5—14 см вод. ст. Монофазные волны повышают внутрикишечное давление в течение 8 с до 30—90 см вод. ст. Медленный компонент сокращений длится от 1 до нескольких минут и повышает давление не столь значительно.
Регуляция моторики тонкой кишки. Моторика тонкой кишки регулируется миогенными, нервными и гуморальными механизмами. Миогенные механизмы обеспечивают автоматию кишечных мышц и сократительную реакцию на растяжение кишки. Однако организованная фазная сократительная деятельность стенки кишки реализуется нейронами мышечно-кишечного миэнтерального (ауэрбахово) нервного сплетения, обладающими ритмической фоновой активностью. Кроме осцилляторов энтеральных метасимпатических узлов имеются два «датчика» ритма кишечных сокращений — первый у места впадения в двенадцатиперстную кишку общего желчного протока, второй — в подвздошной кишке. Деятельность этих «датчиков» и узлов энтерального нервного сплетения контролируется нервными и гуморальными механизмами.
Парасимпатические влияния преимущественно усиливают, симпатические тормозят моторику тонкой кишки. Описаны пептидергические нервные влияния обоих типов. Эффекты раздражения вегетативных нервов в большой мере зависят от состояния кишки, на фоне которого производятся раздражения. Моторику изменяют раздражения спинного и продолговатого мозга, гипоталамуса, лимбической системы, коры больших полушарий. Раздражения ядер передних и средних отделов гипоталамуса преимущественно возбуждают, а заднего — тормозят моторику желудка, тонкой и толстой кишки.
Акт еды тормозит, а затем усиливает кишечную моторику. В дальнейшем она определяется физическими и химическими свойствами химуса: грубая, богатая неперевариваемыми в тонкой кишке пищевыми волокнами и жирами пища ее усиливает.
Местными раздражителями, усиливающими моторику кишки, являются продукты переваривания питательных веществ, особенно жиры, кислоты, щелочи, соли (в концентрированных растворах).
Важное значение для моторики тонкой кишки имеют рефлексы с различных отделов пищеварительного тракта: пищеводно-кишечный (возбуждающий), желудочно-кишечный (возбуждающий и тормозящий), ректоэнтеральный (тормозящий).
|
|
|
Рис. 9.18. Топография всасывания веществ из пищеварительного тракта. |
Дуги этих рефлексов замыкаются на различных уровнях. В целом моторная деятельность любого участка тонкой кишки есть суммарный результат местных, удаленных влияний в пределах пищеварительного тракта (преимущественно возбуждающих влияний с проксимальных и тормозных — с дистальных его отделов относительно раздражаемого участка) и влияний с других систем организма. Гуморальная регуляция. Серотонин, гистамин, гастрин, мотилин, ХЦК, вещество Р, вазопрессин, окситоцин, брадикинин и др., действуя на миоциты или энтеральные нейроны, усиливают, а секретин, ВИП, ГИП и др. тормозят моторику тонкой кишки.
9.6.6. Всасывание различных веществ в тонкой кишке
Всасывание различных веществ приурочено к определенным отделам пищеварительного тракта (рис. 9.18) и осуществляется с помощью различных механизмов (см. раздел 9.3.3.).
Всасывание воды и минеральных солей. Вода поступает в пи щеварительный тракт в составе пищи и выпиваемых жидкостей (2—2,5 л), секретов пищеварительных желез (6—7 л), выводится же с калом 100—150 мл воды. Все остальное количество воды всасывается из пищеварительного тракта в кровь, небольшое количество — в лимфу. Всасывание воды начинается в желудке, но наиболее интенсивно оно происходит в тонкой и особенно толстой кишке (за сутки около 8 л).
Некоторое количество воды всасывается по осмотическому градиенту, хотя вода всасывается и при отсутствии разности осмотического давления. Основное количество воды всасывается из изотонических растворов кишечного химуса, так как в кишечнике гипер- и гипотонические растворы достаточно быстро концентрируются или разводятся. Абсорбция воды из изотонических и гипертонических растворов требует затраты энергии. Активно всасываемые эпителиоцитами растворенные вещества «тянут» за собой воду. Решающая роль в переносе воды принадлежит ионам, особенно Na+, поэтому все факторы, влияющие на его транспорт, изменяют и всасывание воды. Ингибитор натриевого насоса оуабаин подавляет всасывание воды. Всасывание воды сопряжено и с транспортом Сахаров и аминокислот. Так, подавление всасывания Сахаров флорицином замедляет всасывание воды. Многие эффекты замедления или ускорения всасывания воды являются результатом изменения транспорта из тонкой кишки других веществ.
За счет энергии, освобождаемой в тонкой кишке при гликолизе и окислительных процессах, усиливается всасывание воды. Наиболее интенсивно всасывание натрия и воды в кишке осуществляется при рН 6,8 (при рН 3 всасывание воды прекращается).
Изменяют всасывание воды рационы питания. Увеличение в нем доли белка повышает скорость всасывания воды, натрия и хлора.
Скорость всасывания воды изменяется в зависимости от гидра-тированности организма. Наркоз (эфиром и хлороформом), а также ваготомия замедляют всасывание воды. Доказано условно-рефлекторное изменение всасывания воды. На ее всасывание влияют многие гормоны желез внутренней секреции и некоторые гастроинтестинальные гормоны (снижают всасывание воды гастрин, секретин, ХЦК, ВИП, бомбезин, серотонин).
За сутки в желудочно-кишечном тракте всасывается более 1 моля натрия хлорида. У человека натрий почти не всасывается в желудке, интенсивно всасывается в толстой и подвздошной кишке, в тощей кишке его всасывание значительно меньше.
Натрий поступает из полости тонкой кишки в кровь как через кишечные эпителиоциты, так и по межклеточным каналам. Поступление ионов Na+ в эпителиоцит происходит по электрохимическому градиенту пассивным путем. Имеется также система транспорта ионов Na+, сопряженная с транспортом Сахаров и аминокислот, возможно, ионов С1- и НСО3-. Ионы Na+ из эпителиоцитов через их базолатеральные мембраны транспортируются активно в межклеточную жидкость, кровь и лимфу. Это обеспечивает возможность дальнейшего пассивного транспорта ионов Na+ через апикальные мембраны в эпителиоциты из полости кишечника. Различные стимуляторы и ингибиторы всасывания натрия действуют прежде всего на механизмы активного транспорта базолатеральных мембран эпителиоцитов. Интенсивность всасывания натрия зависит от рН кишечного содержимого и гидратации организма.
В толстой кишке всасывание натрия не зависит от наличия Сахаров и аминокислот, а в тонкой кишке зависит. В тонкой кишке переносы ионов Na+ и С1- сопряжены друг с другом, в толстой кишке происходит обмен всасывающегося иона Na+ на ион К+. При снижении содержания в организме натрия его всасывание кишечником резко увеличивается. Усиливают всасывание натрия гормоны гипофиза и надпочечников, угнетают гастрин, секретин и ХЦК.
Всасывание калия происходит в основном в тонкой кишке с помощью механизмов активного и пассивного транспорта по электрохимическому градиенту. Во всасывании калия роль активного транспорта мала, и он сопряжен с транспортом натрия в базолатеральных мембранах эпителиоцитов.
Всасывание ионов хлора происходит в желудке и наиболее активно в подвздошной кишке по типу активного и пассивного транспорта. Пассивный транспорт ионов СUU~ сопряжен с транспортом ионов Na+. Активный транспорт ионов С1- осуществляется через апикальные мембраны, он сопряжен с транспортом ионов Na+ или обменом иона С1- на ион НСОз-
Двузарядные ионы в пищеварительном тракте всасываются очень медленно. Так, в кишечник человека поступает ежесуточно 35 ммоль кальция, но только половина его всасывается. Он всасывается в 50 раз медленнее, чем ион Na , но быстрее, чем двузарядные ионы железа, цинка и марганца. Всасывание кальция совершается с участием переносчиков, активируется желчными кислотами и витамином D, соком поджелудочной железы, некоторыми аминокислотами, натрием, некоторыми антибиотиками. При недостатке кальция в организме его всасывание увеличивается, в чем большую роль могут играть гормоны щитовидной, паращитовидной желез, гипофиза и надпочечников.
Всасывание продуктов гидролиза белков. Белки всасываются в основном в кишечнике после их гидролиза до аминокислот. Всасывание различных аминокислот происходит с неодинаковой скоростью в различных отделах тонкой кишки.
Быстрее других всасываются аргинин, метионин, лейцин, медленнее — фенилаланин, цистеин, тирозин и еще медленнее — аланин, серии, глютаминовая кислота. L-формы аминокислот всасываются интенсивнее, чем D-формы. Всасывание аминокислот из кишки в эпителиоциты через апикальные мембраны осуществляется активно с помощью переносчиков и со значительной затратой энергии фосфорсодержащих макроэргов. Количество аминокислот, всасывающихся пассивно путем диффузии, невелико. В апикальных мембранах эпителиоцитов существует несколько видов переносчиков аминокислот. Из эпителиоцитов аминокислоты транспортируются в межклеточную жидкость по механизму облегченной диффузии. Имеются данные о взаимосвязи транспорта аминокислот через апикальную и базальную мембраны. Большинство аминокислот, образующихся в процессе гидролиза белков и пептидов, всасывается быстрее, чем свободные аминокислоты, введенные в тонкую кишку. Между всасыванием различных аминокислот имеются сложные отношения, в результате которых одни аминокислоты могут ускорять и замедлять всасывание других аминокислот. Транспорт натрия стимулирует всасывание аминокислот. Из менее концентрированных растворов аминокислоты всасываются быстрее, чем из более концентрированных.
Интенсивность всасывания аминокислот зависит от возраста (более интенсивно в молодом возрасте), уровня белкового обмена в организме, содержания в крови свободных аминокислот и ряда других факторов, от нервных и гуморальных влияний.
Всасывание углеводов. Всасывание углеводов происходит в основном в тонкой кишке. С наибольшей скоростью всасываются гексозы, а в их числе глюкоза и галактоза, пентозы всасываются медленнее. Всасывание глюкозы и галактозы осуществляется путем активного транспорта через апикальные мембраны кишечных эпителиоцитов. Они обладают высокой избирательной способностью во всасывании различных углеводов. Транспорт моносахаридов, образующихся при гидролизе олигосахаридов, осуществляется с большей скоростью, чем всасывание моносахаридов, введенных в просвет кишки. Всасывание глюкозы (и некоторых других моносахаридов) через апикальные мембраны кишечных эпителиоцитов активируется транспортом натрия. Глюкоза в отсутствие натрия транспортируется через мембрану в 100 раз медленнее, а против градиента концентрации транспорт глюкозы в этом случае прекращается, что объясняется общностью их переносчика.
Глюкоза аккумулируется в кишечных эпителиоцитах и в последующем транспортируется из них через базолатеральные мембраны в межклеточную жидкость и кровь по градиенту концентрации. Не исключается возможность и активного транспорта их глюкозы через базолатеральные мембраны.
Всасывание фруктозы (и некоторых других моносахаридов) не зависит от транспорта натрия, происходит активно. Не исключают возможность и пассивного транспорта фруктозы через апикальные мембраны эпителиоцитов.
Всасывание углеводов тонкой кишкой усиливается некоторыми аминокислотами, резко тормозится ингибиторами тканевого дыхания.
Различные моносахариды в различных отделах тонкой кишки всасываются с неодинаковой скоростью.
На всасывание Сахаров влияют диета, многие факторы внешней среды, концентрация глюкозы в крови. Существует сложная нервная и гуморальная регуляция всасывания углеводов (Р. О. Файтельберг). Доказано изменение их транспорта под влиянием коры большого мозга, подкорковых структур, ствола головного мозга и спинного мозга. Парасимпатические нервные волокна усиливают, а симпатические тормозят транспорт углеводов из тонкой кишки.
Всасывание глюкозы усиливается гормонами надпочечников, гипофиза, щитовидной железы, а также серотонином, ацетилхолином. Тормозит всасывание глюкозы соматостатин, в меньшей мере — гистамин.
Всасывание продуктов гидролиза липидов. Всасывание различных жиров зависит от их эмульгирования и гидролиза и наиболее активно происходит в двенадцатиперстной кишке и проксимальной части тощей кишки.
В результате действия в полости кишки панкреатической липазы из триглицеридов образуются диглицериды, затем моноглицериды и жирные кислоты, хорошо растворимые в растворах солей желчных кислот. Кишечная липаза производит гидролиз липидов в зоне исчерченной каемки эпителиоцитов. Из моноглицеридов, жирных кислот с участием солей желчных кислот, фосфолипидов и холестерина в полости тонкой кишки образуются мельчайшие мицеллы (диаметром около 100 нм), которые переходят в кишечные эпителиоциты. При этом желчные кислоты мицелл остаются в полости кишки и всасываются в подвздошной кишке по механизму активного транспорта.
В кишечных эпителиоцитах происходит ресинтез триглицеридов. Из них, а также из холестерина, фосфолипидов и глобулинов образуются хиломикроны — мельчайшие жировые частицы, заключенные в тончайшую белковую оболочку. Хиломикроны покидают эпителиоциты через базолатеральные мембраны, переходя в соединительные пространства ворсинок, а оттуда — в центральный лимфатический сосуд ворсинки, чему содействуют ее сокращения. Основное количество жира всасывается в лимфу, поэтому через 3—4 ч после приема пищи лимфатические сосуды наполнены большим количеством лимфы, напоминающей молоко и называемой потому млечным соком.
В нормальных условиях в кровь поступает небольшое количество всосавшегося в кишечнике жира, представленного триглицеридами жирных кислот, молекулы которых содержат короткие углеводородные цепи. В кровеносные капилляры из эпителиоцитов и межклеточного пространства могут транспортироваться и растворимые в воде свободные жирные кислоты и глицерин. В целом же для всасывания жиров, молекулы которых содержат короткие и средние углеводородные цепи, образование в эпителиоцитах хиломикронов необязательно. Небольшое количество хиломикронов может поступать и в кровеносные сосуды ворсинок. Возможно всасывание нейтрального жира в виде молекулярных и мицеллярных растворов.
Скорость гидролиза и всасывание липидов регулируются ЦНС. Парасимпатические нервы ускоряют, а симпатические замедляют всасывание липидов. Стимулируют их всасывание гормоны коркового вещества надпочечников, щитовидной железы и гипофиза, а также гормоны, вырабатываемые в двенадцатиперстной кишке,— секретин и ХЦК.
9.7.1. Поступление кишечного химуса в толстую кишку
Из тонкой кишки химус через илеоцекальный клапан, или сфинктер, — баугиниеву заслонку — порциями переходит в толстую кишку. Сфинктер имеет сложное строение и выполняет роль клапана. Он устроен как губы-складки, суженной частью обращенные в просвет слепой кишки, т. е. подвздошная кишка инвагинирована в слепую. Здесь же сосредоточены циркулярные мышцы сфинктера. Его расслаблению и раскрытию илеоцекального прохода способствуют сокращения продольных мышц тонкой и толстой кишки. При наполнении слепой кишки и ее растяжении сфинктер закрывается и в норме содержимое толстой кишки в тонкую кишку не возвращается.
Вне пищеварения илеоцекальный сфинктер закрыт, но спустя 1—4 мин после приема пищи каждые '/2—1 мин он открывается и химус небольшими порциями (до 15 мл) поступает в толстую кишку. Раскрытие сфинктера происходит рефлекторно: перистальтическая волна тонкой кишки повышает давление в ней и расслабляет илеоцекальный сфинктер и обычно — сфинктер привратника (бисфинктерный рефлекс). Повышение давления в толстой кишке увеличивает тонус илеоцекального сфинктера и тормозит поступление в толстую кишку содержимого тонкой кишки.
За сутки у здорового человека из тонкой в толстую кишку переходит 0,5—4,0 л химуса.
9.7.2. Роль толстой кишки в пищеварении
Пища почти полностью переваривается и всасывается в тонкой кишке. Небольшое количество веществ пищи, в том числе клетчатка и пектин, в составе химуса подвергаются гидролизу в толстой кишке. Гидролиз осуществляется ферментами химуса, микроорганизмов и сока толстой кишки.
Сок толстой кишки в небольшом количестве выделяется вне ее раздражения. Местное механическое раздражение слизистой оболочки увеличивает секрецию в 8—10 раз. Сок состоит из жидкой и плотной частей, имеет щелочную реакцию (рН 8,5—9,0). Плотную часть сока составляют слизистые комочки из отторгнутых кишечных эпителиоцитов и слизи, секретируемой бокаловидными клетками.
Основное количество ферментов содержится в плотной части сока; их активность значительно меньше, чем в тонкой кишке, хотя спектры ферментов близки. В соке толстой кишки нет энтерокиназы и сахаразы, щелочной фосфатазы содержится в 15—20 раз меньше, чем в соке тонкой кишки. В соке толстой кишки содержится небольшое количество катепсина, пептидазы, липазы, амилазы и нуклеазы.
С участием этих ферментов в проксимальной части толстой кишки происходит гидролиз питательных веществ. В зависимости от осмотического и гидростатического давления кишечного содержимого интенсивно всасывается вода (до 4—6 л за сутки). Химус постепенно превращается в каловые массы (за сутки выводится 150—250 г сформированного кала). При употреблении растительной пищи их больше, чем после приема смешанной или мясной пищи. Если пища богата неперевариваемыми волокнами (целлюлоза, гемицеллюлоза, пектин, лигнин), то количество кала увеличивается не только за счет них, но и вследствие ускорения передвижения химуса и формируемого кала, что предотвращает запоры и их патогенные последствия.
9.7.3. Моторная функция толстой кишки
Весь процесс пищеварения у взрослого человека длится 1— 3 сут, из них наибольшее время приходится на пребывание остатков пищи в толстой кишке. Ее моторика обеспечивает резервуарную функцию — накопление содержимого, всасывание из него ряда веществ, в основном воды, продвижение его, формирование каловых масс и их удаление (дефекация).
У здорового человека контрастная масса через 3—З'/2 ч после ее приема начинает поступать в толстую кишку, которая заполняется в течение 24 ч и полностью опорожняется за 48—72 ч.
Содержимое слепой кишки совершает небольшие и длительные перемещения то в одну, то в другую сторону за счет медленных сокращений кишки. Для толстой кишки характерны сокращения нескольких типов: малые и большие маятникообразные, перистальтические и антиперистальтические, пропульсивные. Первые четыре типа сокращений обеспечивают перемешивание содержимого кишки и повышение давления в ее полости, что способствует сгущению содержимого путем всасывания воды. Сильные пропульсивные сокращения возникают 3—4 раза в сутки и продвигают кишечное содержимое в дистальном направлении.
Толстая кишка имеет интра- и экстрамуральную иннервацию, играющую ту же роль, что и у тонкой кишки. Толстая кишка получает парасимпатическую иннервацию в составе блуждающих и тазовых нервов; парасимпатические влияния усиливают моторику путем условных и безусловных рефлексов при раздражении пищевода, желудка и тонкой кишки. Симпатические нервы проходят в составе чревных нервов и тормозят моторику кишки.
Ведущее значение в организации моторики толстой кишки имеют интрамуральные нервные механизмы при местном механическом и химическом раздражении толстой кишки ее содержимым. Раздражение механорецепторов прямой кишки тормозит моторику вышележащих отделов тонкой кишки. Тормозят ее и серотонин, адреналин, глюкагон.
9.7.3. Дефекация
Дефекация — опорожнение толстой кишки от каловых масс наступает в результате раздражения рецепторов прямой кишки накопившимися в ней каловыми массами. Позыв на дефекацию возникает при повышении давления в прямой кишке до 40—50 см вод. ст. Давление 20—30 см вод. ст. вызывает чувство наполнения прямой кишки. Сфинктеры прямой кишки — внутренний, состоящий из гладких мышц, и наружный, образованный поперечнополосатой мускулатурой, вне дефекации находятся в состоянии тонического сокращения. В результате рефлекторного расслабления этих сфинктеров, перистальтических сокращений кишки, сокращения мышцы, поднимающей задний проход (m. levator any), укорачивающей дистальную часть прямой кишки, сокращений ее кольцевых мышц кал выбрасывается из прямой кишки. В этом большое значение имеет так называемое натуживание, при котором сокращаются мышцы брюшной стенки и диафрагмы, повышается внут-рибрюшное давление, достигающее при акте дефекации 220 см вод. ст. Первичная рефлекторная дуга от рецепторов прямой кишки замыкается в пояснично-крестцовом отделе спинного мозга (рис. 9.19). Эта рефлекторная дуга обеспечивает непроизвольный акт дефекации. Произвольный акт осуществляется при участии коры больших полушарий мозга, центров продолговатого мозга и гипоталамуса.
Из спинального центра дефекации по парасимпатическим нервным волокнам в составе тазового нерва поступают импульсы, тормозящие тонус сфинктеров и усиливающие моторику прямой кишки, стимулируя акт дефекации. Симпатические нервные влияния повышают тонус сфинктеров и тормозят моторику прямой кишки.
Произвольный компонент акта дефекации состоит в нисходящих влияниях головного мозга на спинальный центр, в расслаблении наружного сфинктера, сокращении диафрагмы и брюшных мышц. У большинства здоровых людей акт дефекаций совершается 1—2 раза в сутки.
Газы толстой кишки. За сутки из кишечника при дефекации и вне ее выводится 100—500 мл газа. При метеоризме объем его может достигать Зли более. Растяжение толстой кишки газом вызывает состояние дискомфорта, чувство распирания. Растяжение газом тонкой кишки вызывает болевые ощущения.
Газы кишечника имеют различное происхождение. Часть их попадает в пищеварительный тракт при заглатывании пищи, и у страдающих аэрофагией в желудке и кишечнике содержится повышенное количество газа. В основном газ образуется в кишечнике.
|
|
|
Рис. 9.19. Рефлекторное обеспечение дефекации. Р — рецепторы; 1 — внутренний сфинктер заднего прохода; 2 — наружный сфинктер заднего прохода; 3 — анальные железы; 4 — интрамуральные парасимпатические ганглии; (+) —влияния, повышающие тонус мышцы; (—) — влияния, понижающие тонус мышцы. |
При взаимодействии гидрокарбонатов секрета поджелудочной железы с кислыми продуктами кишечного химуса образуется значительное количество СО2. Газы продуцируются и микрофлорой кишечника. При переваривании некоторых видов пищи при участии микрофлоры образуется большое количество газов (бобы, капуста, лук, черный хлеб, картофель и др.).
У здоровых людей покидающую кишечник газовую смесь составляют N2 (24—90%), СО2 (4,3—29%), О2 (0,1—23%), Н2 (0,6—47%), метан (0—26%), в небольшом количестве сероводород, аммиак, меркаптан.
9.8. МИКРОФЛОРА ПИЩЕВАРИТЕЛЬНОГО ТРАКТА
Пищеварительный тракт человека и животных «заселен» микроорганизмами. В одних отделах тракта в норме их содержание незначительно или они почти отсутствуют, в других их находится очень много. Макроорганизм и его микрофлора составляют единую динамичную экологическую систему. Динамичность эндоэко-логического микробного биоценоза пищеварительного тракта определяется количеством поступающих в него микроорганизмов (у человека за сутки перорально поступает около 1 млрд микробов), интенсивностью их размножения и гибели в пищеварительном тракте и выведения из него микробов в составе кала (у человека в норме выделяется за сутки 1012—1014 микроорганизмов).
Каждый из отделов пищеварительного тракта имеет характерные для него количество и набор микроорганизмов. Их число в полости рта, несмотря на бактерицидные свойства слюны, велико (107—108 клеток на 1 мл ротовой жидкости). Содержимое желудка здорового человека натощак благодаря бактерицидным свойствам желудочного сока часто бывает стерильным, но нередко обнаруживается и относительно большое число микроорганизмов (до 103 на 1 мл содержимого), проглатываемых со слюной. Примерно такое же количество их в двенадцатиперстной и начальной части тощей кишки. В содержимом подвздошной кишки микроорганизмы обнаруживаются регулярно, и число их в среднем составляет 106 на 1 мл содержимого. В содержимом толстой кишки число бактерий максимальное, и 1 г кала здорового человека содержит 10 млрд и более микроорганизмов.
Микрофлору кишечника делят на три группы: 1-я — главная; в ее состав входят бифидобактерии и бактероиды, которые составляют 90% от всех микробов; 2 — сопутствующая (лактобактерии, эшерихии, энтерококки, 10% от общего числа микроорганизмов); 3 — остаточная (цитробактер, энтеробактер, протеи, дрожжи, клостридии, стафилококки, аэробные бациллы и др., менее 1%). Анаэробная микрофлора преобладает над аэробной.
Микроорганизмы, связанные со слизистой оболочкой кишечника, относятся к мукозной микрофлоре — М-микрофлоре, а локализованные в полости кишки — к полостной — П-микрофлоре. Соотношения между М- и П-микрофлорой динамичны, определяются многими факторами. К внешним воздействиям М-микрофлора более устойчива, чем П-микрофлора.
За илеоцекальным клапаном (баугиниева заслонка) резко изменяется не только число, но и качество микрофлоры. Толстая кишка является своеобразной микроэкологической зоной. В ней П-микрофлора представлена бактероидами, бифидобактериями, лактобактериями, вейлонеллами, клостридиями, пептострептококками, пептококками, энтеробактериями, аэробными бациллами, дифтероидами, энтерококками, стафилококками, микрококками, плесневыми грибами (преобладают бактероиды, бифидобактерии, лактобактерии). М-микрофлора слизистой оболочки толстой кишки отличается от микрофлоры полости кишки, и в М-микрофлоре наибольшее число бифидо- и лактобактерии. Общее число М-форм слизистой оболочки толстой кишки составляет у людей 106, с соотношением анаэробов к аэробам 10:1. Максимальное число бактерий в фекалиях (1010—1013 на 1 г), где они составляют до 30 % от их массы. В качественном соотношении она сходна с микрофлорой полости толстой кишки.
Состав и количество микроорганизмов в пищеварительном тракте зависит от эндогенных и экзогенных факторов. К первым относятся влияния слизистой оболочки пищеварительного канала, его секретов, моторики и самих микроорганизмов. Ко вторым — характер питания, факторы внешней среды, прием антибактериальных препаратов. Экзогенные факторы влияют непосредственно и опосредованно через эндогенные факторы. Например, прием той или иной пищи изменяет секреторную и моторную деятельность пищеварительного тракта, что трансформирует его микрофлору.
Существенны влияния на микрофлору функционального состояния пищеварительной системы. Перистальтика пищеварительного тракта обеспечивает транспорт микроорганизмов в составе химуса в дистальном направлении, что играет определенную роль в создании проксимодистального градиента заселенности кишечника микроорганизмами. Дискинезии кишечника изменяют этот градиент.
Баугиниева заслонка, играющая роль клапана, а также более высокое давление содержимого перед заслонкой, чем за ней, предотвращают поступление микроорганизмов с содержимым из толстой кишки в тонкую.
В формировании микрофлоры пищеварительного тракта велика роль пищеварительных секретов. Слюна имеет муромидазу (лизо-цим), которая определяет бактерицидные свойства этого секрета. Желудочный сок за счет соляной кислоты и других факторов обладает бактерицидностью, что существенно влияет на численность и состав микрофлоры кишечника. Количество и состав микрофлоры зависят и от поступления в тонкую кишку сока поджелудочной железы, кишечного секрета и желчи. Эти влияния не только прямые, но и опосредованные. Так, снижение содержания питательных веществ в химусе лишает микроорганизмы необходимых им питательных веществ. Гидролизаты белков и жиров ингибируют развитие ряда микроорганизмов. Такое действие оказывают лейкодиапедез и свободные желчные кислоты, а также выделяемые в составе пищеварительных секретов не только лизоцим, но и иммуноглобулины, С-рективный белок, лактоферрин.
Важным экзогенным фактором является питание. Его стабильность, сбалансированность и адекватность важны в стабилизации эубиоза человека. Вегетарианская диета способствует увеличению количества энтерококков и эубактерий. Избыточный прием животных белков и жиров вызывает повышение в составе микрофлоры числа клостридий, бактероидов. Избыток в рационе животных жиров ведет к увеличению числа бактероидов и уменьшению числа бифидобактерий и энтерококков; недостаток жиров вызывает обратные изменения микрофлоры. Молочная диета (лактоза) способствует повышению количества бифидобактерий.
|
|
|
Рис. 9.20. Потоки веществ из пищеварительного тракта во внутреннюю среду организма (по А. М. Уголеву). |
Включение в рацион человека нефизиологических компонентов (например, белки одноклеточных), сублимированных продуктов неблагоприятно влияет на микрофлору кишечника.
Нормальная микрофлора — эубиоз — выполняет ряд важнейших для макроорганизма функций. Исключительно важным является ее участие в формировании иммунобиологической реактивности организма. Эубиоз предохраняет макроорганизм от внедрения и размножения в нем патогенных микроорганизмов. Нарушение нормальной микрофлоры при заболевании или в результате длительного введения антибактериальных препаратов нередко влечет за собой осложнения, вызываемые бурным размножением в кишечнике дрожжей, стафилококка, протея и других микроорганизмов.
Кишечная микрофлора синтезирует витамины К и группы В, которые частично покрывают потребность в них. Микрофлора синтезирует и другие вещества, важные для организма.
Ферменты бактерий расщепляют не переваренные в тонкой кишке целлюлозу, гемицеллюлозу и пектины, и образовавшиеся продукты используются макроорганизмом. У разных людей количество целлюлозы, гидролизуемое ферментами бактерий, неодинаковое и составляет в среднем около 40%. Гемицеллюлоза метаболизируется в большей мере, чем целлюлоза.
Микроорганизмы утилизируют непереваренные пищевые вещества, образуя при этом ряд веществ, которые всасываются из кишечника и включаются в обмен веществ организма. Микрофлора существенно влияет на печеночно-кишечную циркуляцию компонентов желчи и через них — на деятельность печени. С участием микрофлоры кишечника в организме происходит обмен белков, фосфолипидов, желчных и жирных кислот, билирубина, холестерина.
Пищеварительные секреты, выполнив свою физиологическую роль, частично разрушаются и всасываются в тонкой кишке, а частично поступают в толстую кишку. Здесь они подвергаются действию микрофлоры. Микроорганизмы принимают участие в разложении парных желчных кислот, ряда органических веществ с образованием органических кислот, их аммонийных солей, аминов и др.
Итак, из пищи реально формируется не два потока: всасываемые питательные вещества (нутриенты) и невсасываемый балласт. На самом деле потоков веществ больше (рис. 9.20): кроме первичных нутриентов, образовавшихся в результате гидролиза питательных веществ пищи, существует значительный поток вторичных нутриентов. По А. М. Уголеву, это три потока: 1-й — модифицированные микрофлорой нутриенты; 2-й — продукты жизнедеятельности бактерий; 3-й — модифицированные балластные вещества.
К числу таких веществ относятся не только упомянутые выше витамины, но и другие физиологически активные амины (кадаверин, октамин, терамин, пиперидин, диметиламин, гистамин и др.), незаменимые аминокислоты, углеводы, жиры.
Существенны и небактериальные потоки эндогенных веществ, в их числе инкретируемые гормоны и ферменты, а также образовавшиеся из пептидов пищи в результате их гидролиза экзогормоны. Среди потоков всасываемых веществ и токсичные продукты деятельности кишечной микрофлоры.
Анатомическое положение печени на пути крови, несущей питательные и иные вещества от пищеварительного тракта, особенности строения, кровоснабжения, лимфообращения, специфика функций гепатоцитов определяют функции этого органа. Ранее описана желчеотделительная функция печени, но она не единственная.
Важна также барьерная функция печени, состоящая в обезвреживании токсичных соединений, поступивших с пищей либо образовавшихся в кишечнике за счет деятельности его микрофлоры, лекарств, всосавшихся в кровь и принесенных кровью к печени. Химические вещества обезвреживаются путем их ферментативного окисления, восстановления, метилирования, ацетилирования, гидролиза (1-я фаза) и последующей конъюгации с рядом веществ (глюкуроновой, серной и уксусной кислотами, глицином, таурином и др. — 2-я фаза). Не все вещества обезвреживаются в две фазы: некоторые — в одну или без изменений выводятся в составе желчи и мочи, особенно растворимые конъюгаты. Нейтрализация токсичного аммиака происходит за счет образования мочевины и креатинина. Микроорганизмы обезвреживаются в основном путем фагоцитоза и лизиса их.
Печень принимает участие в инактивации ряда гормонов (глюкокортикоиды, альдостерон, андрогены, эстрогены, инсулин, глюкагон, ряд гастроинтестинальных гормонов) и биогенных аминов (гистамин, серотонин, катехоламины).
Экскреторная функция печени выражается в выделении из крови в составе желчи большого числа веществ, обычно трансформированных в печени, что является ее участием в обеспечении гомеостаза.
Печень участвует в обмене белков: в ней синтезируются белки крови (весь фибриноген, 95% альбуминов, 85% глобулинов), происходят дезаминирование и переаминирование аминокислот, образование мочевины, глутамина, креатина, факторов свертывания крови и фибринолиза (1, II, V, VII, IX, X, XII, XIII, антитромбин, антиплазмин). Желчные кислоты влияют на транспортные свойства белков крови.
Печень участвует в обмене липидов: в их гидролизе и всасывании, синтезе триглицеридов, фосфолипидов, холестерина, желчных кислот, липопротеидов, ацетоновых тел, окислении триглицеридов. Велика роль печени в обмене углеводов: здесь осуществляются процессы гликогенеза, гликогенолиза, включение в обмен глюкозы, галактозы и фруктозы, образование глюкуроновой кислоты.
Печень участвует в эритрокинетике, в том числе в разрушении эритроцитов, деградации гема с последующим образованием билирубина.
Важна роль печени в обмене витаминов (особенно жирорастворимых A, D, Е, К), всасывание которых в кишечнике происходит с участием желчи. Ряд витаминов депонируется в печени и высвобождается по мере их метаболической потребности (A, D, К, С, РР). Депонируются в печени микроэлементы (железо, медь, марганец, кобальт, молибден и др.) и электролиты. Печень участвует в иммунопоэзе и иммунологических реакциях.
Выше упоминалась кишечно-печеночная циркуляция желчных кислот. Важно их участие не только в гидролизе и всасывании липидов, но и в других процессах. Желчные кислоты являются регуляторами холереза и выделения в составе желчи холестерина, желчных пигментов, активности печеночных цитоферментов, влияют на транспортную активность энтероцитов, ресинтез в них триглицеридов, регулируют пролиферацию, передвижение и отторжение энтероцитов с кишечных ворсинок.
Регуляторное влияние желчи распространяется на секрецию желудка, поджелудочной железы и тонкой кишки, эвакуаторную деятельность гастродуоденального комплекса, моторику кишечника, реактивность органов пищеварения к нейротрансмиттерам, регуляторным пептидам и аминам.
Циркулирующие с кровью желчные кислоты влияют на многие физиологические процессы: при повышении концентрации желчных кислот в крови физиологические процессы угнетаются — в этом и проявляется токсическое действие желчных кислот; нормальное их содержание в крови поддерживает и стимулирует физиологические и биохимические процессы.
9.10. НЕПИЩЕВАРИТЕЛЬНЫЕ ФУНКЦИИ ПИЩЕВАРИТЕЛЬНОГО ТРАКТА
Все физиологические системы и органы имеют основные и дополнительные функции. В большой мере это присуще и пищеварительной системе.
9.10.1. Экскреторная деятельность пищеварительного тракта
Пищеварительные железы и кишечник выводят из крови в ходе секреции и путем рекреции многие эндогенные и экзогенные вещества, участвуя таким образом в сохранении гомеостаза организма. Так, экзосекреция железами желудка Н+ и HC03- поджелудочной железой HCO33 имеет существенное значение в поддержании постоянства кислотно-основного состояния организма. Путем выделения в полость пищеварительного тракта метаболитов (первая группа выводимых веществ) организм особождается от них (например, выделение в составе желудочного сока мочевины). Вторая группа веществ выводится из крови и депонируется в содержимом пищеварительного тракта (например, вода и растворенные в ней неорганические соли). Третья группа выделенных с секретом в химус веществ подвергается гидролизу, всасывается и включается в метаболизм (например, белки в количестве 60 г в сутки, что немаловажно для эндогенного питания). Четвертая группа веществ этих трансформаций не претерпевает, но участвует в пищеварительной деятельности и циркулирует между кровотоком и содержимым пищеварительного тракта (например, кишечно-печеночная циркуляция желчных кислот).
В пищеварительный тракт выводятся и экзогенные вещества: ряд лекарственных, токсичных веществ, попавших в кровоток энтеральным и парентеральным путем.
9.10.2. Участие пищеварительного тракта в водно-солевом обмене
Пищеварительный тракт участвует в нескольких этапах данного обмена. Это участие просматривается уже в формировании чувства жажды в результате неприятного ощущения сухости во рту, которое снижается при слюноотделении. В свою очередь оно зависит от количества воды в организме.
Доказаны орофарингеальный, желудочный и кишечный сенсорные механизмы возбуждения и торможения центра жажды с пищеварительного тракта. Дегидратация (обезвоживание) организма снижает секреторную активность пищеварительных желез, что способствует сохранению воды в организме. Диурез и объем секреции, выделение электролитов в составе секретов желез и мочи взаимосвязаны и также направлены на сохранение воды в организме.
В одних случаях этот процесс одно-, в других случаях — разнонаправленный.
Значительное количество воды и электролитов депонируется в пищеварительном тракте и включается в их обмен, циркулирует между кровью и содержимым пищеварительного тракта. Ряд регуляторных пептидов пищеварительного тракта влияет на водно-солевой обмен (см. раздел 10.1.4).
9.10.2. Эндокринная функция пищеварительного тракта и выделение в составе секретов биологически активных веществ
Регуляторные пептиды пищеварительного тракта влияют не только на секрецию, моторику, всасывание, высвобождение других регуляторных пептидов и пролиферацию органов пищеварения, но оказывают и так называемые общие эффекты. Они многочисленны и проявляются в ряде поведенческих реакций, но особенно выражены в изменении обмена веществ, деятельности сердечно-сосудистой и эндокринной систем организма.
Приведем примеры общих эффектов регуляторных пептидов пищеварительного тракта. Гастрин усиливает высвобождение гистамина, инсулина, кальцитонина, липолиз в жировой ткани, выделение почками воды, калия, натрия. Соматостатин тормозит высвобождение гастроинтестинальных гормонов, соматотропина, подавляет гликогенолиз, изменяет пищевое поведение. ВИП снижает тонус кровеносных сосудов с гипотензивным эффектом, тонус бронхов. Малые дозы ВИП вызывают гипертермию. Секретин усиливает липолиз и гликолиз, тормозит реабсорбцию гидрокарбонатов в почках, увеличивает диурез, ренальное выделение натрия и калия, повышает сердечный выброс. ХЦК является рилизинг-фактором для инсулина. /7/7 угнетает аппетит. ГИП усиливает высвобождение инсулина и глюкагона. Нейротензин усиливает высвобождение глюкагона, соматостатина, вазопрессина, гистамина, лютеинизирующего и фолликулостимулирующего гормонов, тормозит высвобождение инсулина, усиливает теплопродукцию.
С учетом того, что двенадцатиперстная кишка является «гипофизом брюшной полости» (А. М. Уголев), становится понятным возникновение широкого спектра нарушений обмена веществ при удалении в экспериментах двенадцатиперстной кишки, при ее клинической патологии и дуоденэктомии.
Ряд регуляторных пептидов образуется из экзогенных (в том числе пищевых) белков при их частичном переваривании, ограниченном протеолизе, в желудке и кишечнике. Так образуется группа морфиноподобных веществ (экзорфины) при гидролизе белков молока и хлеба.
На высвобождение регуляторных пептидов и аминов пищеварительного тракта влияют гормоны эндокринных желез, т. е. между этими эндокринными системами имеется двусторонняя связь, деятельность этих эндокринных систем контролируется и нервно-рефлекторными механизмами.
Не менее сложным является вопрос о содержании в секретах пищеварительных желез и в ткани желез многих биологически активных веществ. Они могут синтезироваться в самих железах, элиминироваться из крови и затем выделяться в составе секретов. Слюна содержит лизоцим (муромидазу), который обладает антибактериальной активностью, участвует в реакциях местного иммунитета, увеличивая продукцию антител, фагоцитов, повышает межклеточную проницаемость (подобно гиалуронидазе), свертываемость крови. Калликреин слюны и слюнных желез принимает участие в образовании эндогенных вазодилататоров и гипотензивных веществ, участвует в обеспечении местной гиперемии и повышении проницаемости капилляров, усиливает секрецию желудка.
Из слюны выделен белок, обладающий свойствами антианемического фактора. Ферменты слюны влияют на микрофлору полости рта, на трофику ее слизистой оболочки и зубов. Слюнные железы принимают участие в обеспечении гомеостаза ферментов и гормонов в крови, выделяя их из крови и в кровь.
Одним из многих обнаруженных в слюне и железе веществ является паротин. Он влияет на обмен белков, кальция (увеличивает кальцификацию трубчатых костей и зубов), липидов, гемопоэз, пролиферацию хрящевой ткани, увеличивает васкуляризацию органов, проницаемость гистогематических барьеров, сперматогенез.
В железе найдены факторы стимуляции роста нервов и эпителия, дающие многочисленные эффекты. Накоплены факты о связи слюнных желез с активностью щитовидной, паращитовидной желез, гипофизом, надпочечниками, поджелудочной железой, тимусом.
Желудок выполняет многие не пищеварительные функции. Его сок обладает высокой бактерицидностью, содержит антианемический фактор Касла (транскоррин), про-, антикоагулянты и фибринолитики. В желудке образуется ряд регуляторных пептидов и аминов широкого спектра физиологической активности.
Секрет поджелудочной железы принимает участие в регуляции микрофлоры кишечника, трофики его слизистой оболочки и скорости обновления ее эпителиоцитов. Хроническая потеря сока поджелудочной железы вызывает глубокие нарушения углеводного, жирового, белкового и водно-солевого обмена, деятельности кроветворных органов и некоторых эндокринных желез. Эти нарушения временно купируются подкожным введением нативного панкреатического секрета, что свидетельствует о содержании в нем важных веществ. Велика роль в обмене веществ эндокринного аппарата поджелудочной железы (см. раздел 5.2.6). Железа образует ряд регуляторных пептидов (ВИП, гастрин, энкефалин, ПП) и ферментов (в том числе калликреин), липоксин — «гормон жирового обмена», ваготонин, повышающий тонус парасимпатической части автономной нервной системы.
Кишечник участвует во многих видах обмена и гомеостаза, содержит и выделяет многие регуляторные пептиды. Слизистая оболочка тонкой кишки обладает тромбопластичесхой, антигепариновой и фибринолитической активностью.
Многочисленные проявления патологии пищеварительной системы обусловлены не только нарушением пищеварительных функций и ассимиляции пищи, но и важных не пищеварительных функций этой системы.
9.10.3. Инкреция (эндосекреция) пищеварительными железами ферментов
Установлена прямая связь между небольшой гидролитической активностью крови и мочи и секретов пищеварительных желез. После частичного или полного удаления желудка или поджелудочной железы активность продуцируемых ими ферментов и концентрация их предшественников в крови и моче резко понижалась: нарушение оттока секрета из желез вызывала противоположный эффект. Эти открытия привлекли внимание клиницистов, так как давали возможность беззондового исследования функционального состояния пищеварительных желез с диагностической целью. Этот метод получил широкое распространение, однако физиологическое значение инкреции ферментов исследовано недостаточно.
Ферменты, синтезируемые пищеварительными железами, транспортируются в лимфу и кровь из интерстициальной жидкости, куда попадают инкреторным путем непосредственно из гландулоцитов, покидая их через базолатеральные мембраны; резорбируются из протоков желез и из тонкой кишки, высвобождаются из разрушенных гландулоцитов.
Чем больше секреторных клеток, продуцирующих данный фермент, тем выше показатели его экзосекреции в составе сока и инкреции — содержание и активность в крови, ренальное и экстраренальное выделение этого фермента. При повышении сопротивления оттоку секрета из железы (обтурация протока, отек слизистой оболочки желудка, повышение давления в полости, куда выводится секрет) экзосекреция снижается, но возрастает транспорт ферментов в кровь. Стимуляция секреции желез на эндосекреции ферментов отражается в меньшей мере, чем на их экзосекреции.
Относительное постоянство активности пищеварительных ферментов в периферической крови является результатом сбалансированности количества поступающих в крови ферментов с количеством ферментов, подвергающихся катаболизму, ренальному и экстраренальному выведению из организма. Наиболее изучено выделение некоторых ферментов в составе мочи (ренальное выделение). Ферменты выделяются также с потом, молоком, рекретируются ферменты и в составе секретов пищеварительных желез (экстраренальное выделение), включаются в пищеварительный процесс и подвергаются аутолизу, гидролизу протеазами секретов.
Инкретированные ферменты в крови находятся в свободном и связанном с транспортными белками и форменными элементами состояниях. Из крови ферменты адсорбируются эндотелием кровеносных сосудов. Есть свидетельства участия инкретированных ферментов в гидролизе пищевых веществ крови и лимфы, т. е. эти ферменты включены в метаболизм всего организма. Ферменты крови выполняют и регуляторную роль — тормозят секрецию одноименных ферментов, но могут усиливать секрецию других ферментов данной железы. Так, показано стимулирующее влияние трипсиногена крови на секрецию пепсиногена железами желудка, пепсиногена крови — на секрецию трипсиногена поджелудочной железой. Экзо- и эндосекретируемые ферменты выполняют роль гидролаз и модуляторов (модулирующие эффекты вызывают также их фрагменты) секреторной и моторной деятельности пищеварительных органов. Пускового влияния на них в физиологических дозах ферменты не оказывают.
9.10.4. Иммунная система пищеварительного тракта
Пищеварительный тракт имеет ряд защитных механизмов против патогенных антигенных факторов. Среди них уже назывались антибактериальные свойства слюны, сока поджелудочной железы, желчи, протеолитическая активность секретов, моторная деятельность кишечника, характерная ультраструктура поверхности слизистой оболочки тонкой кишки, препятствующая проникновению через нее бактерий. К этим неспецифическим барьерным механизмам следует добавить специфическую иммунную систему защиты, локализованную в пищеварительном тракте и составляющую важную часть общей многокомпонентной иммунной системы человека.
В пищеварительном тракте имеется три группы иммунокомпетентных элементов лимфоидной ткани:
1) лимфоидные фолликулы на всем протяжении пищеварительного тракта; в подвздошной кишке и червеобразном отростке эти фолликулы образуют большие скопления в виде групповых лимфоидных узелков (пейеровы бляшки);
2) плазматические и Т-лимфоидные клетки слизистой оболочки пищеварительного тракта;
3) малые неидентифицированные лимфоидные клетки.
К органам местной иммунной системы пищеварительного тракта, где локализованы эти элементы, относятся миндалины глоточного кольца в устье дыхательного и пищеварительного трактов; солитарные лимфатические фолликулы, расположенные в стенке кишки на всем ее протяжении, крупные лимфоидные образования — пейеровы бляшки в наибольшем количестве расположены в подвздошной кишке, встречаются в двенадцатиперстной и тощей кишке; червеобразный отросток; плазматические клетки слизистой оболочки желудка и кишечника. Местная иммунная система пищеварительного тракта обеспечивает две основные функции: 1) распознавание и индукцию толерантности к пищевым антигенам; 2) блокирующий эффект по отношению к патогенным микроорганизмам.
Миндалины осуществляют местную защиту путем выделения в полость глотки иммуноглобулинов, интерферона, лизоцима, лимфоцитов, макрофагов и простагландинов. Они способствуют формированию иммунной памяти путем образования клона лимфоцитов, которые подготавливают иммунную систему к повторной встрече с антигенами.
Групповые лимфоидные узелки, или пейеровы бляшки, являясь иммунокомпетентными элементами тонкой кишки, участвуют в распознавании пищевых антигенов химуса и формировании местного иммунного ответа. Червеобразный отросток является важнейшим компонентом местной иммунной системы. Вследствие поступления в него антигенов пищевого и микробного происхождения развивается иммунный ответ.
Плазматические клетки свободно располагаются в слизистой оболочке и строме ворсинок кишки под эпителием. Они синтезируют и секретируют иммуноглобулины всех известных в настоящее время классов (G, M, A, D, Е). Различные отделы пищеварительного тракта в норме содержат различное количество плазмоцитов, продуцирующих соответственно разное количество иммуноглобулинов (Ig) разных классов с преобладанием IgA. При напряженном иммуногенезе его секреция многократно возрастает. Секреторный иммуноглобулин A (sIgA) — особая форма данного класса глобулинов; он не разрушается протеолитическими пищеварительными ферментами. Нормальная кишечная микрофлора человека имеет большое значение в развитии секреторной иммунной системы и особенно в синтезе и секреции sIgA. Он находится на апикальной части эпителиальных клеток и в межэпителиальном пространстве, на поверхности слизистых оболочек, в смеси с секретированной слизью. Она вместе с антигенами и микроорганизмами удаляется в полость пищеварительного тракта. Способность отталкиваться от эпителиальных клеток проявляется у sIgA после соединения его с антигеном.
sIgA осуществляет защитную функцию, непосредственно действуя на бактерии, связывая их и препятствуя внедрению в глубь слизистой оболочки. sIgA дезактивирует токсичные продукты деятельности бактерий, небактериальные продукты и способствует последующему их разрушению протеолитическими ферментами.
Органы местной иммунной защиты пищеварительного тракта обеспечивают защитный иммунный ответ на контакт с антигенами, перорально поступающими в организм человека из внешней среды. Выраженность ответной реакции зависит не только от силы антигенной стимуляции, но и от функционального состояния макроорганизма, его нервной и эндокринной регуляторных систем, в том числе и от влияний регуляторных пептидов пищеварительного тракта. При всей своей автономности местная иммунная система пищеварительного тракта тем не менее состоит в сложной связи с общей иммунной системой и другими местными иммунными системами человека.